

Ovarian hormones are not required for PRL-induced mammary tumorigenesis, but estrogen enhances neoplastic processes
- PMID: 19635758
- PMCID: PMC2841967
- DOI: 10.1677/JOE-09-0221
Ovarian hormones are not required for PRL-induced mammary tumorigenesis, but estrogen enhances neoplastic processes
Abstract
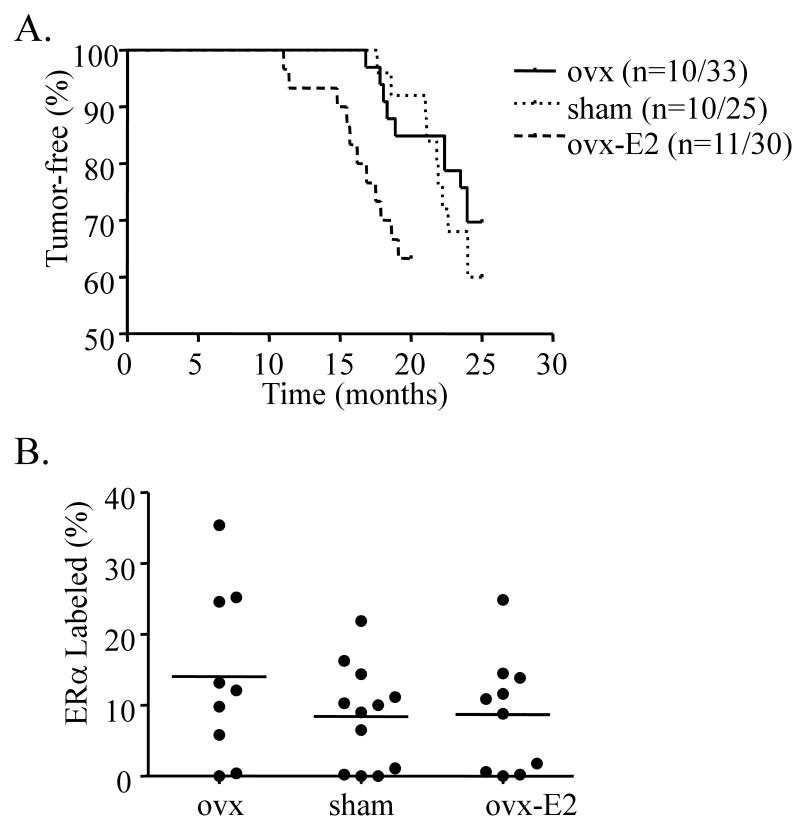
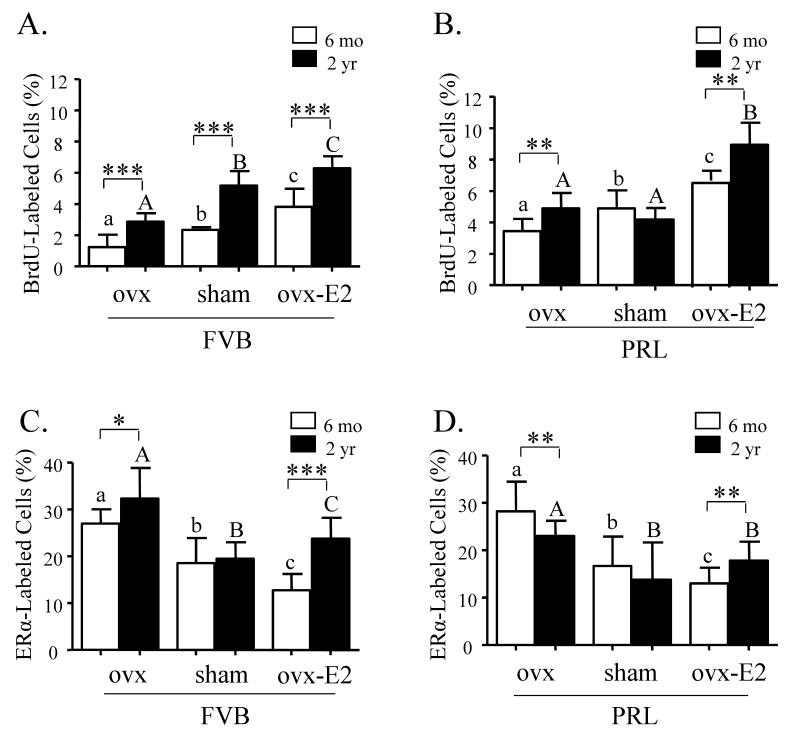
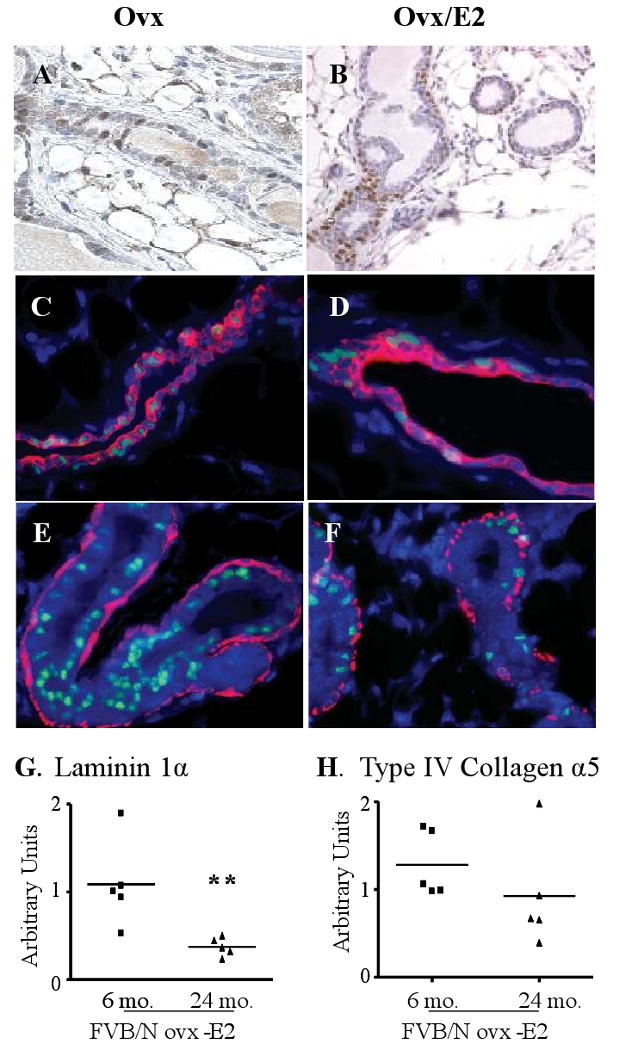
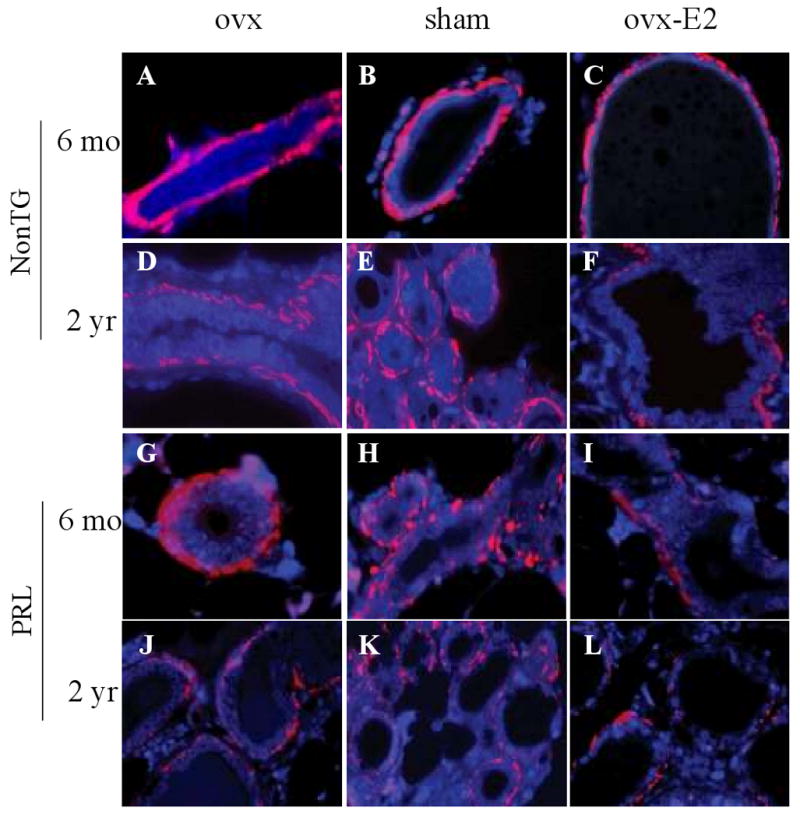
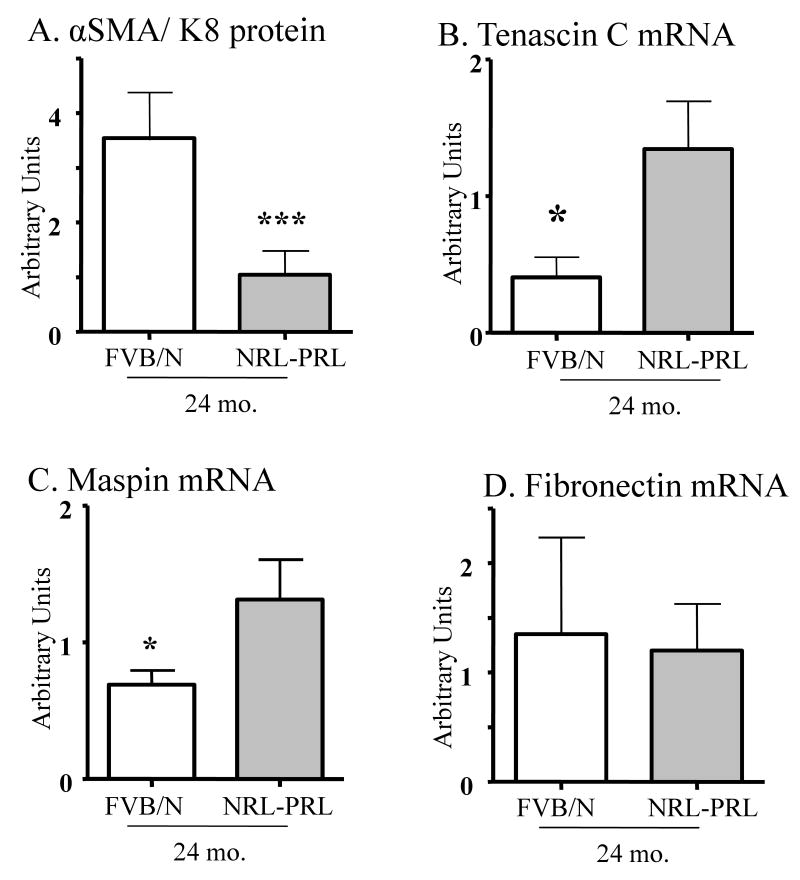
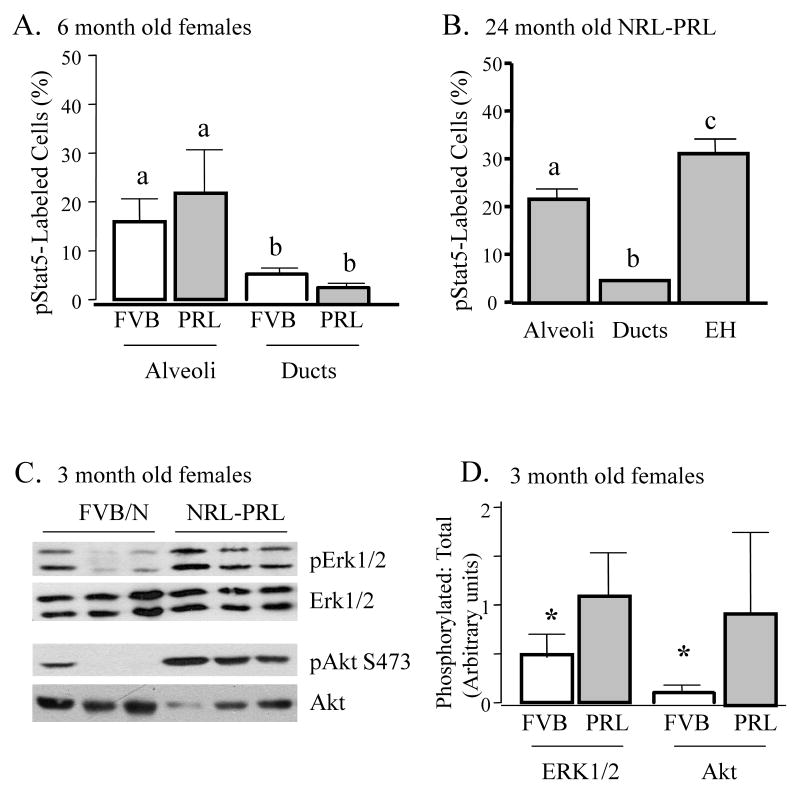
Epidemiologic studies have demonstrated that increased prolactin (PRL) exposure raises the risk of invasive estrogen receptor alpha (ERalpha)-positive breast cancer in women. However, the mechanism(s) whereby this occurs and the interactions with estrogen itself in this disease remain poorly understood. In order to investigate the role of ovarian hormones in the disease process, we employed a transgenic model neu-related lipocalin (NRL)-PRL in which transgenic PRL is directed to mammary epithelial cells by the PRL- and estrogen-insensitive NRL promoter, mimicking the endogenous PRL expression within the breast observed in women. This high local exposure leads to mammary lesion development and eventually carcinomas. Ovariectomy (ovx), shortly after puberty, did not alter the incidence or latency of PRL-induced mammary carcinomas, consistent with the independence of PRL from circulating estrogens as a risk factor for invasive breast cancer in women. However, chronic estrogen administration to ovx NRL-PRL females decreased the latency of both ERalpha-positive and -negative tumors. We identified multiple mechanisms that may underlie this observation. Elevated estrogen exposure cooperated with PRL to increase epithelial proliferation and myoepithelial abnormalities, increasing the incidence of preneoplastic lesions. Critical components of the extracellular matrix secreted by the myoepithelium were reduced with age, and transgenic PRL raised transcripts for tenascin-C and maspin, both associated with tumor progression and poor prognosis in subclasses of clinical breast tumors. Mammary pERK1/2 and pAkt, but not phosphorylated Stat5, were markedly elevated by local PRL. Together, these findings indicate that PRL employs multiple mechanisms to promote mammary tumorigenesis.
Conflict of interest statement
Figures
References
-
- Alarid ET. Lives and times of nuclear receptors. Molecular Endocrinology. 2006;20:1972–1981. - PubMed
-
- Anderson E, Clarke RB, Howell A. Estrogen receptor in mammary gland physiology. In: Ethier SP, editor. Contemporary Endocrinology: Endocrine Oncology. Totowa, NJ: Humana Press Inc; 2000. pp. 1–16.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
