

E-prostanoid 3 receptor deletion improves pulmonary host defense and protects mice from death in severe Streptococcus pneumoniae infection
- PMID: 19635910
- PMCID: PMC4016992
- DOI: 10.4049/jimmunol.0900129
E-prostanoid 3 receptor deletion improves pulmonary host defense and protects mice from death in severe Streptococcus pneumoniae infection
Abstract
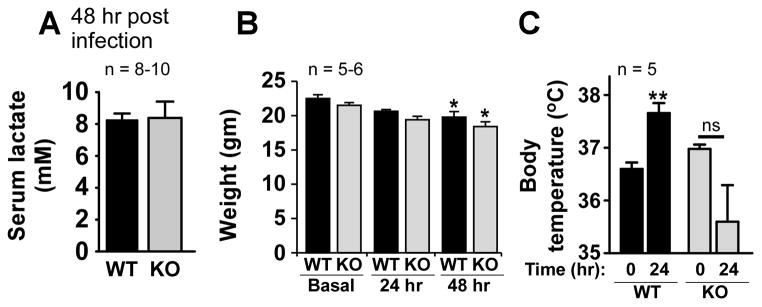
Prostaglandins (PGs) are potent lipid mediators that are produced during infections and whose synthesis and signaling networks present potential pharmacologic targets for immunomodulation. PGE(2) acts through the ligation of four distinct G protein-coupled receptors, E-prostanoid (EP) 1-4. Previous in vitro and in vivo studies demonstrated that the activation of the G(alphas)-coupled EP2 and EP4 receptors suppresses inflammatory responses to microbial pathogens through cAMP-dependent signaling cascades. Although it is speculated that PGE(2) signaling via the G(alphai)-coupled EP3 receptor might counteract EP2/EP4 immunosuppression in the context of bacterial infection (or severe inflammation), this has not previously been tested in vivo. To address this, we infected wild-type (EP3(+/+)) and EP3(-/-) mice with the important respiratory pathogen Streptococcus pneumoniae or injected mice i.p. with LPS. Unexpectedly, we observed that EP3(-/-) mice were protected from mortality after infection or LPS. The enhanced survival observed in the infected EP3(-/-) mice correlated with enhanced pulmonary clearance of bacteria; reduced accumulation of lung neutrophils; lower numbers of circulating blood leukocytes; and an impaired febrile response to infection. In vitro studies revealed improved alveolar macrophage phagocytic and bactericidal capacities in EP3(-/-) cells that were associated with an increased capacity to generate NO in response to immune stimulation. Our studies underscore the complex nature of PGE(2) immunomodulation in the context of host-microbial interactions in the lung. Pharmacological targeting of the PGE(2)-EP3 axis represents a novel area warranting greater investigative interest in the prevention and/or treatment of infectious diseases.
Figures






References
-
- Statistics, N. C. f. H; U. S. D. o. H. a. H. Services, editor. Health, United States, 2007, with chartbook on trends in the health of Americans. U.S. Government Printing Office; Hyattsville, MD: 2007. p. 567.
-
- Mandell LA, Wunderink RG, Anzueto A, Bartlett JG, Campbell GD, Dean NC, Dowell SF, File TM, Jr, Musher DM, Niederman MS, Torres A, Whitney CG. Infectious Diseases Society of America/American Thoracic Society consensus guidelines on the management of community-acquired pneumonia in adults. Clin Infect Dis. 2007;44(Suppl 2):S27–72. - PMC - PubMed
-
- Levine OS, O’Brien KL, Knoll M, Adegbola RA, Black S, Cherian T, Dagan R, Goldblatt D, Grange A, Greenwood B, Hennessy T, Klugman KP, Madhi SA, Mulholland K, Nohynek H, Santosham M, Saha SK, Scott JA, Sow S, Whitney CG, Cutts F. Pneumococcal vaccination in developing countries. Lancet. 2006;367:1880–1882. - PubMed
-
- Kadioglu A, Andrew PW. The innate immune response to pneumococcal lung infection: the untold story. Trends Immunol. 2004;25:143–149. - PubMed
-
- Aronoff DM, Canetti C, Peters-Golden M. Prostaglandin E2 inhibits alveolar macrophage phagocytosis through an E-prostanoid 2 receptor-mediated increase in intracellular cyclic AMP. J Immunol. 2004;173:559–565. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
