

Structural models of human eEF1A1 and eEF1A2 reveal two distinct surface clusters of sequence variation and potential differences in phosphorylation
- PMID: 19636410
- PMCID: PMC2712093
- DOI: 10.1371/journal.pone.0006315
Structural models of human eEF1A1 and eEF1A2 reveal two distinct surface clusters of sequence variation and potential differences in phosphorylation
Abstract
Background: Despite sharing 92% sequence identity, paralogous human translation elongation factor 1 alpha-1 (eEF1A1) and elongation factor 1 alpha-2 (eEF1A2) have different but overlapping functional profiles. This may reflect the differential requirements of the cell-types in which they are expressed and is consistent with complex roles for these proteins that extend beyond delivery of tRNA to the ribosome.
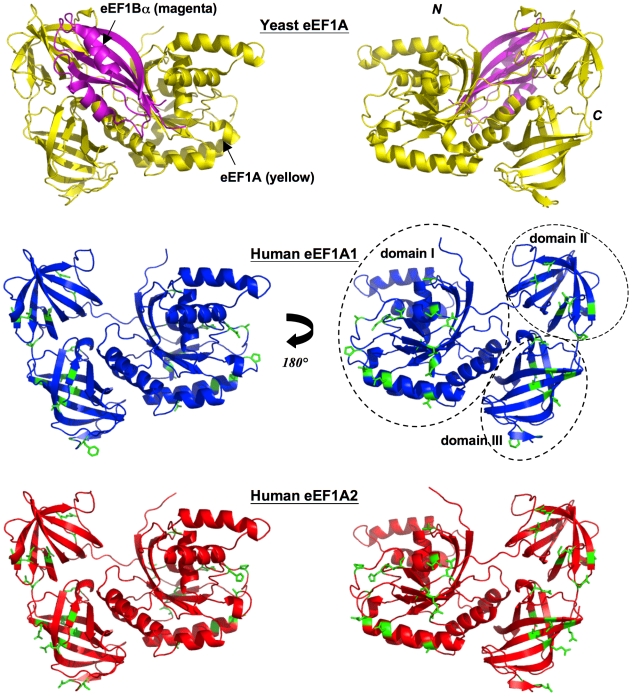
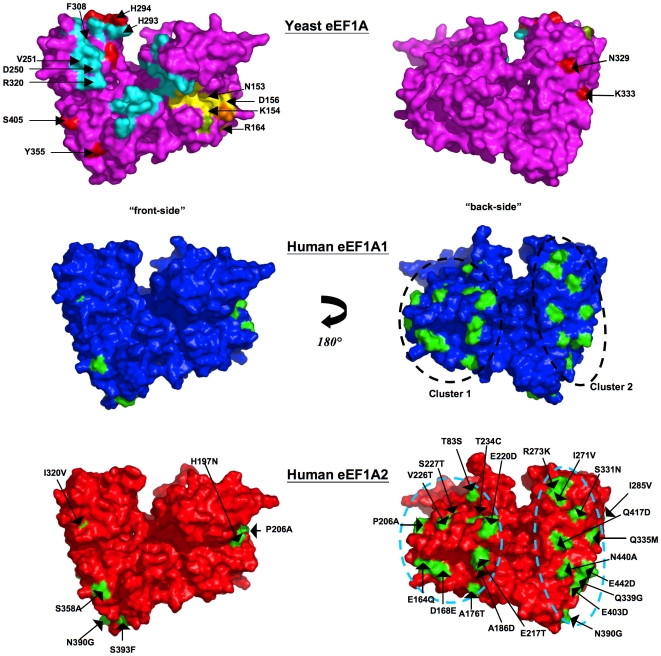
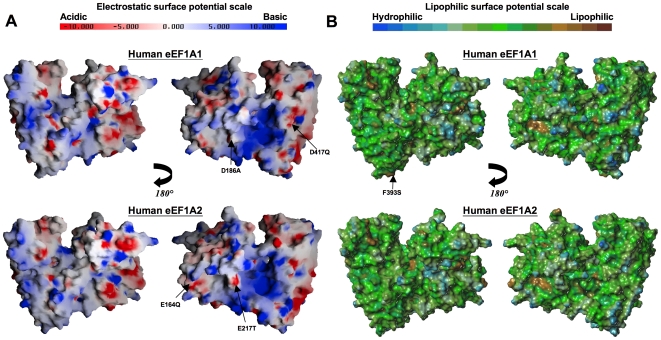
Methodology/principal findings: To investigate the structural basis of these functional differences, we created and validated comparative three-dimensional (3-D) models of eEF1A1 and eEF1A2 on the basis of the crystal structure of homologous eEF1A from yeast. The spatial location of amino acid residues that vary between the two proteins was thereby pinpointed, and their surface electrostatic and lipophilic properties were compared. None of the variations amongst buried amino acid residues are judged likely to have a major structural effect on the protein fold, or to affect domain-domain interactions. Nearly all the variant surface-exposed amino acid residues lie on one face of the protein, in two proximal but distinct sub-clusters. The result of previously performed mutagenesis in yeast may be interpreted as confirming the importance of one of these clusters in actin-bundling and filament disorganization. Interestingly, some variant residues lie in close proximity to, and in a few cases show differences in interactions with, residues previously inferred to be directly involved in binding GTP/GDP, eEF1Balpha and aminoacyl-tRNA. Additional sequence-based predictions, in conjunction with the 3-D models, reveal likely differences in phosphorylation sites that could reconcile some of the functional differences between the two proteins.
Conclusions: The revelation and putative functional assignment of two distinct sub-clusters on the surface of the protein models should enable rational site-directed mutagenesis, including homologous reverse-substitution experiments, to map surface binding patches onto these proteins. The predicted variant-specific phosphorylation sites also provide a basis for experimental verification by mutagenesis. The models provide a structural framework for interpretation of the resulting functional analysis.
Conflict of interest statement
Figures


References
-
- Lund A, Knudsen SM, Vissing H, Clark B, Tommerup N. Assignment of human elongation factor 1alpha genes: EEF1A maps to chromosome 6q14 and EEF1A2 to 20q13.3. Genomics. 1996;36:359–361. - PubMed
-
- Lee S, Francoeur AM, Liu S, Wang E. Tissue-specific expression in mammalian brain, heart, and muscle of S1, a member of the elongation factor-1 alpha gene family. J Biol Chem. 1992;267:24064–24068. - PubMed
-
- Newbery HJ, Loh DH, O'Donoghue JE, Tomlinson VA, Chau YY, et al. Translation elongation factor eEF1A2 is essential for post-weaning survival in mice. J Biol Chem. 2007;282:28951–28959. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
