

Crossover inhibition in the retina: circuitry that compensates for nonlinear rectifying synaptic transmission
- PMID: 19636690
- PMCID: PMC2766457
- DOI: 10.1007/s10827-009-0170-6
Crossover inhibition in the retina: circuitry that compensates for nonlinear rectifying synaptic transmission
Abstract
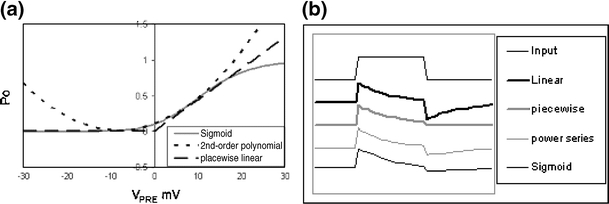
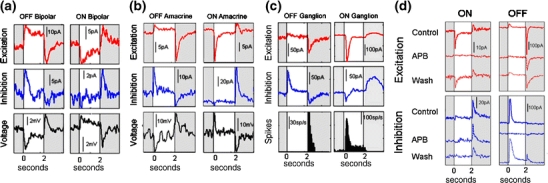
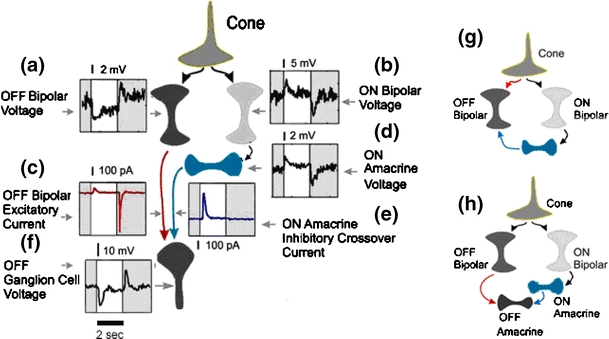
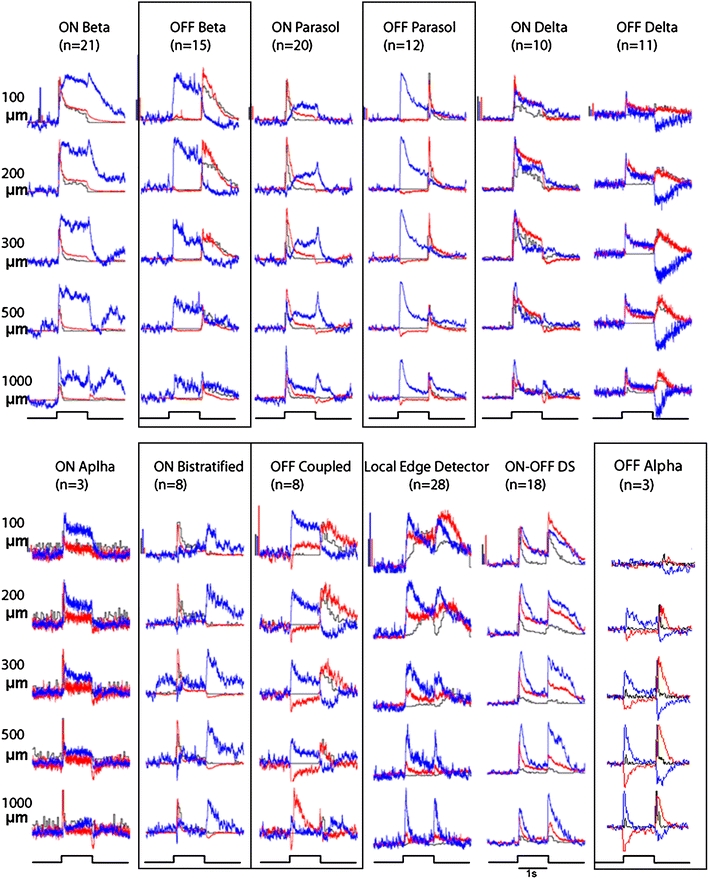
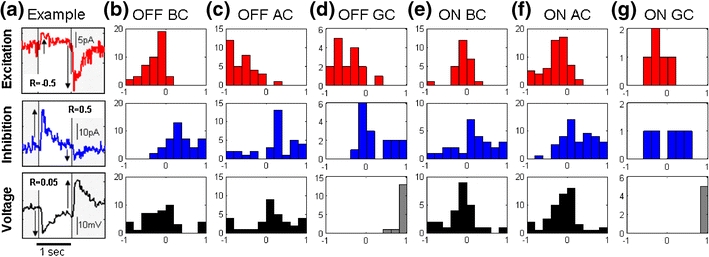
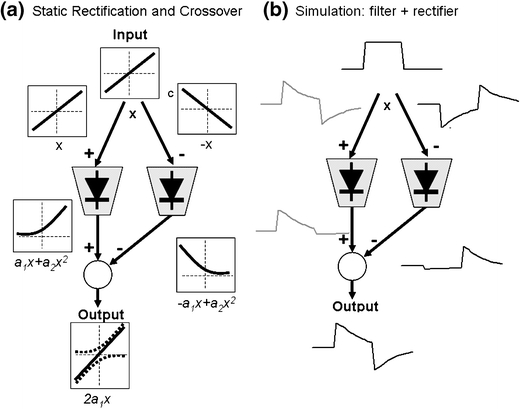
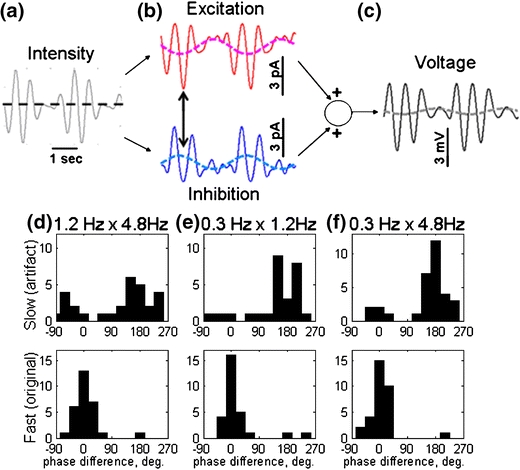
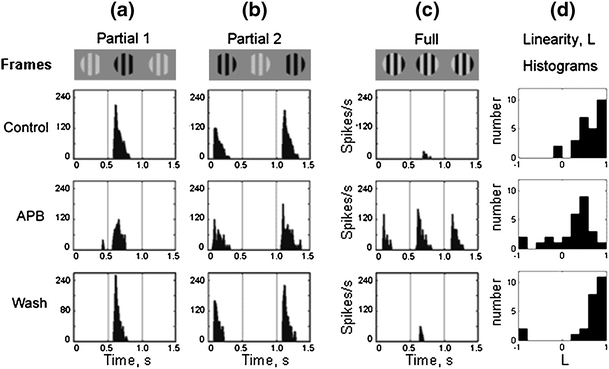
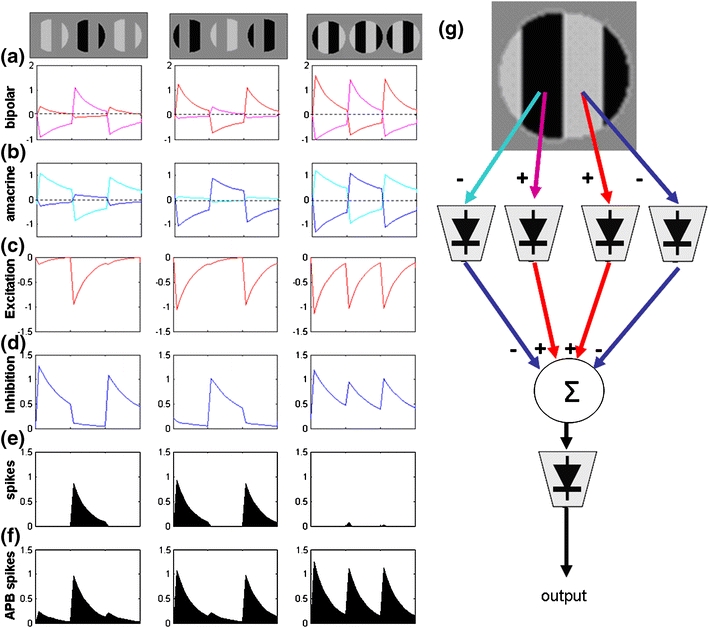
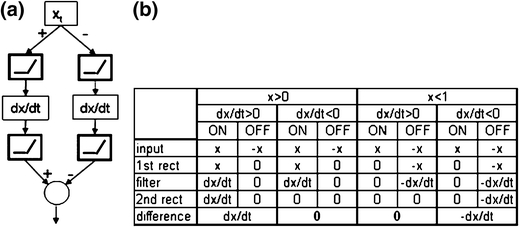
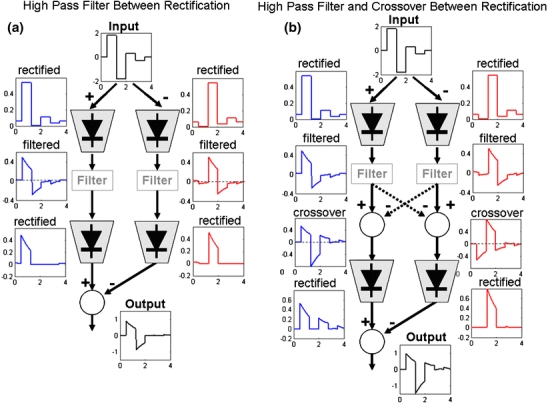
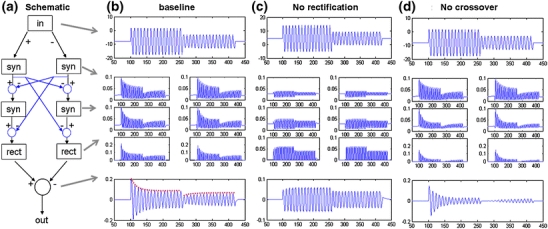
In the mammalian retina, complementary ON and OFF visual streams are formed at the bipolar cell dendrites, then carried to amacrine and ganglion cells via nonlinear excitatory synapses from bipolar cells. Bipolar, amacrine and ganglion cells also receive a nonlinear inhibitory input from amacrine cells. The most common form of such inhibition crosses over from the opposite visual stream: Amacrine cells carry ON inhibition to the OFF cells and carry OFF inhibition to the ON cells ("crossover inhibition"). Although these synapses are predominantly nonlinear, linear signal processing is required for computing many properties of the visual world such as average intensity across a receptive field. Linear signaling is also necessary for maintaining the distinction between brightness and contrast. It has long been known that a subset of retinal outputs provide exactly this sort of linear representation of the world; we show here that rectifying (nonlinear) synaptic currents, when combined thorough crossover inhibition can generate this linear signaling. Using simple mathematical models we show that for a large set of cases, repeated rounds of synaptic rectification without crossover inhibition can destroy information carried by those synapses. A similar circuit motif is employed in the electronics industry to compensate for transistor nonlinearities in analog circuits.
Figures


References
-
- Anderson JS, Carandini M, Ferster D. Orientation tuning of input conductance, excitation, and inhibition in cat primary visual cortex. Journal of Neurophysiology. 2000;84:909–926. - PubMed
