

Insig-mediated, sterol-accelerated degradation of the membrane domain of hamster 3-hydroxy-3-methylglutaryl-coenzyme A reductase in insect cells
- PMID: 19638338
- PMCID: PMC2785366
- DOI: 10.1074/jbc.M109.032342
Insig-mediated, sterol-accelerated degradation of the membrane domain of hamster 3-hydroxy-3-methylglutaryl-coenzyme A reductase in insect cells
Abstract
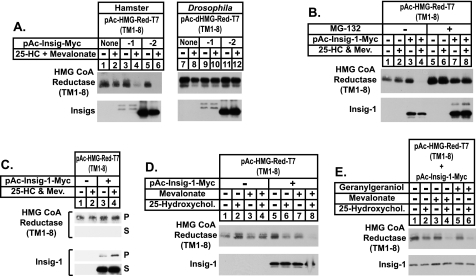
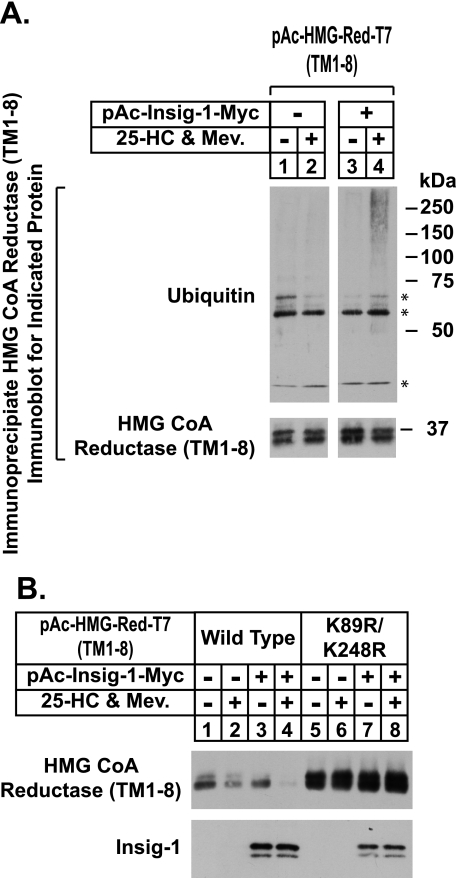
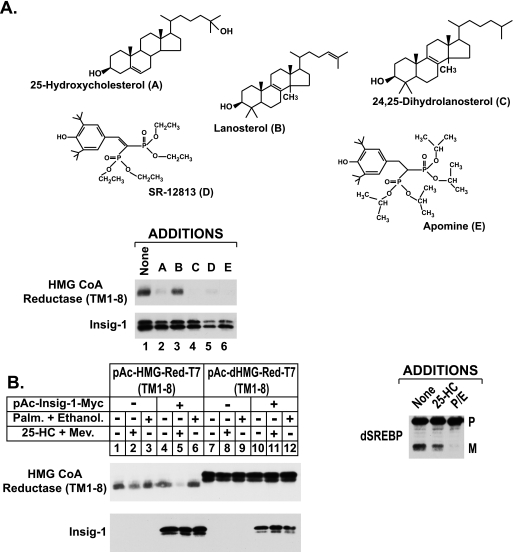
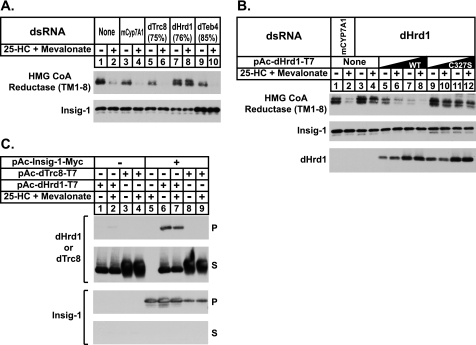
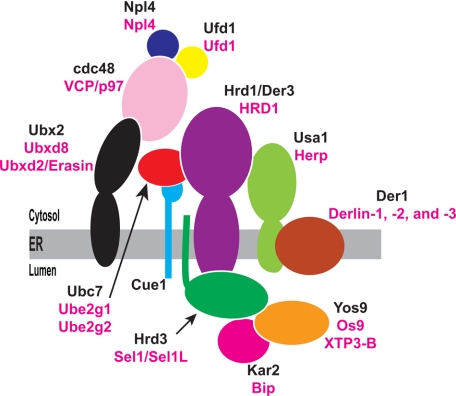
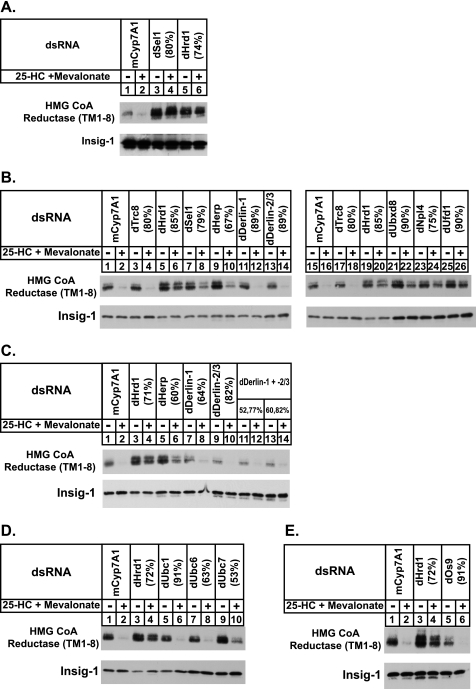
Sterol-accelerated degradation of the enzyme 3-hydroxy-3-methylglutaryl-coenzyme A reductase is one of several mechanisms through which cholesterol synthesis is controlled in mammalian cells. This degradation results from sterol-induced binding of the membrane domain of reductase to endoplasmic reticulum membrane proteins called Insig-1 and Insig-2, which are carriers of a ubiquitin ligase called gp78. The ensuing gp78-mediated ubiquitination of reductase is a prerequisite for its rapid, 26 S proteasome-mediated degradation from endoplasmic reticulum membranes, a reaction that slows a rate-limiting step in cholesterol synthesis. Here, we report that the membrane domain of hamster reductase is subject to sterol-accelerated degradation in Drosophila S2 cells, but only when mammalian Insig-1 or Insig-2 are co-expressed. This degradation mimics the reaction that occurs in mammalian cells with regard to its absolute requirement for the action of Insigs, sensitivity to proteasome inhibition, augmentation by nonsterol isoprenoids, and sterol specificity. RNA interference studies reveal that this degradation requires the Drosophila Hrd1 ubiquitin ligase and several other proteins, including a putative substrate selector, which associate with the enzyme in yeast and mammalian systems. These studies define Insigs as the minimal requirement for sterol-accelerated degradation of the membrane domain of reductase in Drosophila S2 cells.
Figures
References
-
- Goldstein J. L., Brown M. S. (1990) Nature 343, 425–430 - PubMed
-
- Gil G., Faust J. R., Chin D. J., Goldstein J. L., Brown M. S. (1985) Cell 41, 249–258 - PubMed
-
- Liscum L., Finer-Moore J., Stroud R. M., Luskey K. L., Brown M. S., Goldstein J. L. (1985) J. Biol. Chem. 260, 522–530 - PubMed
-
- Sever N., Yang T., Brown M. S., Goldstein J. L., DeBose-Boyd R. A. (2003) Mol. Cell 11, 25–33 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
