

The dynamic properties of intermediate filaments during organelle transport
- PMID: 19638410
- PMCID: PMC2724608
- DOI: 10.1242/jcs.046789
The dynamic properties of intermediate filaments during organelle transport
Abstract
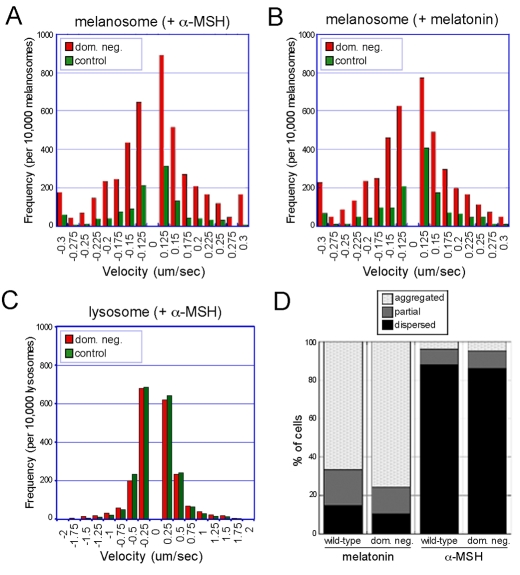
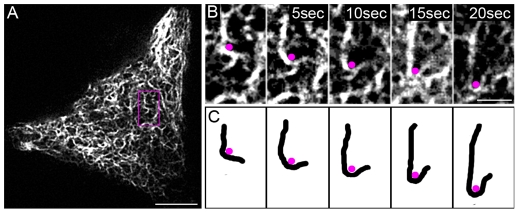
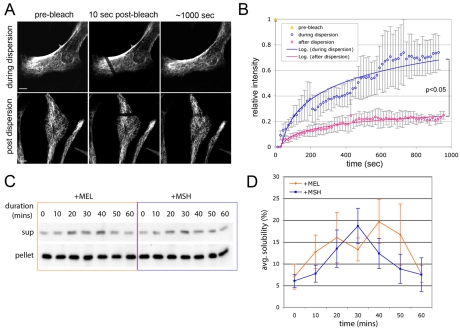
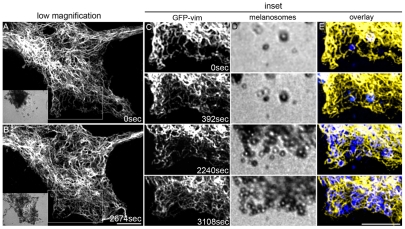
Intermediate filament (IF) dynamics during organelle transport and their role in organelle movement were studied using Xenopus laevis melanophores. In these cells, pigment granules (melanosomes) move along microtubules and microfilaments, toward and away from the cell periphery in response to alpha-melanocyte stimulating hormone (alpha-MSH) and melatonin, respectively. In this study we show that melanophores possess a complex network of vimentin IFs which interact with melanosomes. IFs form an intricate, honeycomb-like network that form cages surrounding individual and small clusters of melanosomes, both when they are aggregated and dispersed. Purified melanosome preparations contain a substantial amount of vimentin, suggesting that melanosomes bind to IFs. Analyses of individual melanosome movements in cells with disrupted IF networks show increased movement of granules in both anterograde and retrograde directions, further supporting the notion of a melanosome-IF interaction. Live imaging reveals that IFs, in turn, become highly flexible as melanosomes disperse in response to alpha-MSH. During the height of dispersion there is a marked increase in the rate of fluorescence recovery after photobleaching of GFP-vimentin IFs and an increase in vimentin solubility. These results reveal a dynamic interaction between membrane bound pigment granules and IFs and suggest a role for IFs as modulators of granule movement.
Figures
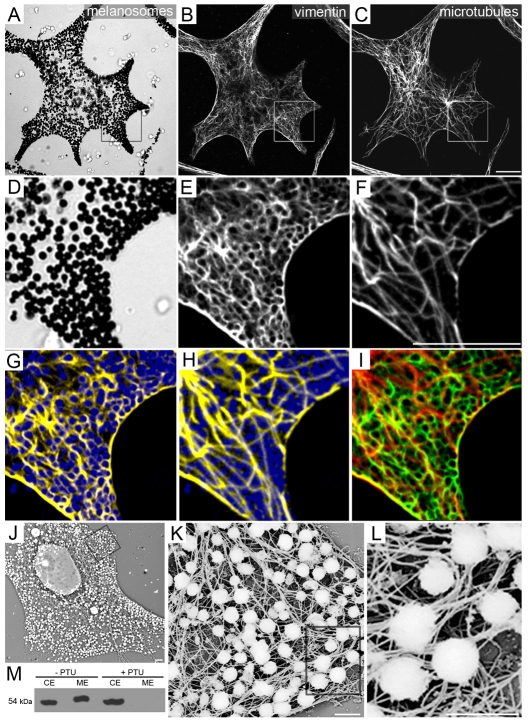
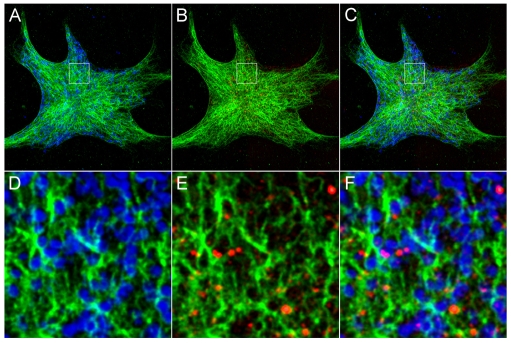
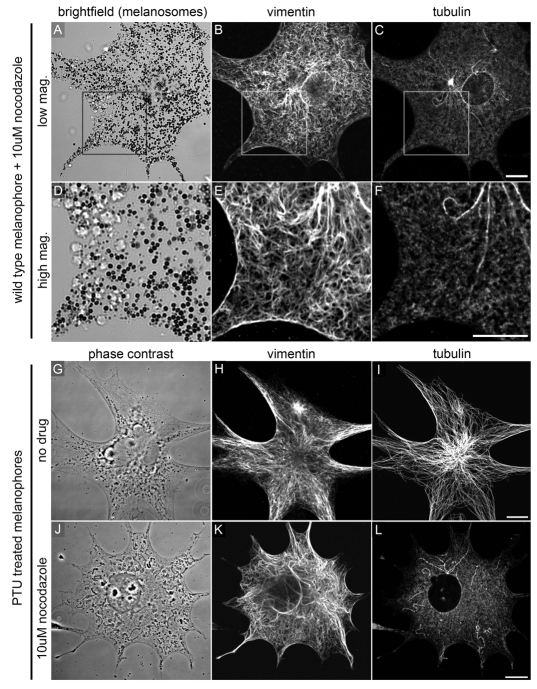
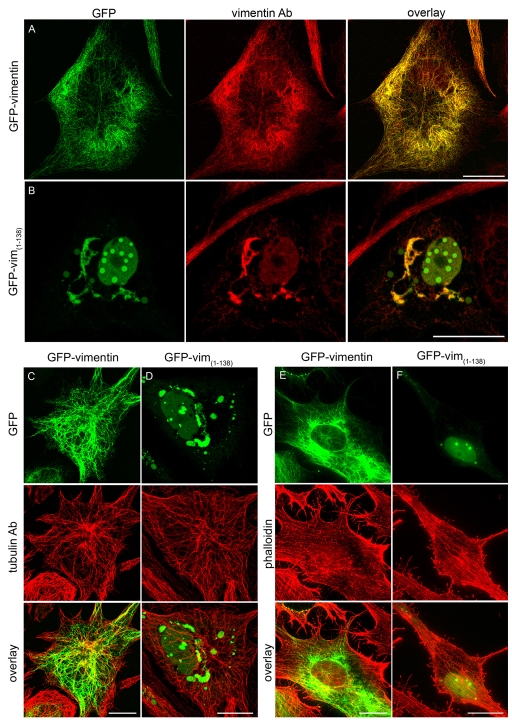
References
-
- Chang, L. and Goldman, R. D. (2004). Intermediate filaments mediate cytoskeletal crosstalk. Nat. Rev. Mol. Cell Biol. 5, 601-613. - PubMed
-
- Chijiwa, T., Mishima, A., Hagiwara, M., Sano, M., Hayashi, K., Inoue, T., Naito, K., Toshioka, T. and Hidaka, H. (1990). Inhibition of forskolin-induced neurite outgrowth and protein phosphorylation by a newly synthesized selective inhibitor of cyclic AMP-dependent protein kinase, N-[2-(p-bromocinnamylamino)ethyl]-5-isoquinolinesulfonamide (H-89), of PC12D pheochromocytoma cells. J. Biol. Chem. 265, 5267-5272. - PubMed
-
- Chou, Y. H., Flitney, F. W., Chang, L., Mendez, M., Grin, B. and Goldman, R. D. (2007). The motility and dynamic properties of intermediate filaments and their constituent proteins. Exp. Cell Res. 313, 2236-2243. - PubMed
-
- Collier, N. C., Sheetz, M. P. and Schlesinger, M. J. (1993). Concomitant changes in mitochondria and intermediate filaments during heat shock and recovery of chicken embryo fibroblasts. J. Cell Biochem. 52, 297-307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
