

A structural basis for the pH-dependent xanthophyll cycle in Arabidopsis thaliana
- PMID: 19638474
- PMCID: PMC2729612
- DOI: 10.1105/tpc.109.068007
A structural basis for the pH-dependent xanthophyll cycle in Arabidopsis thaliana
Abstract
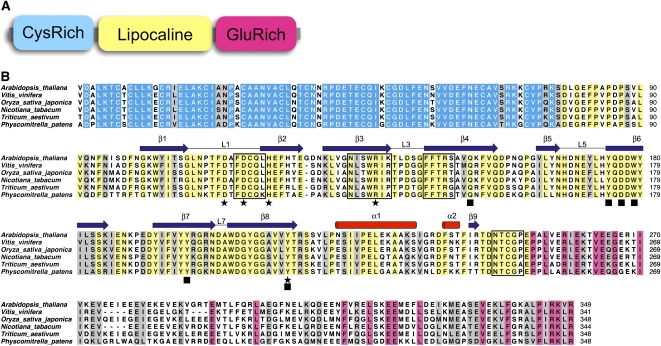
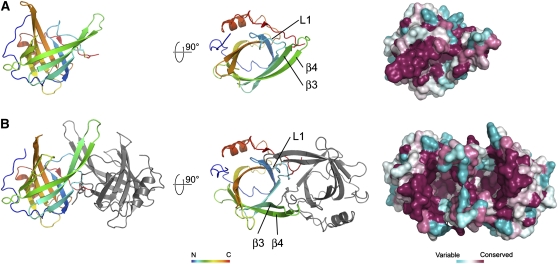
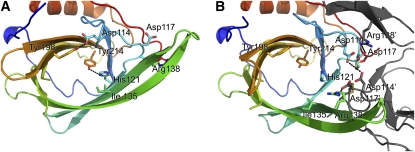
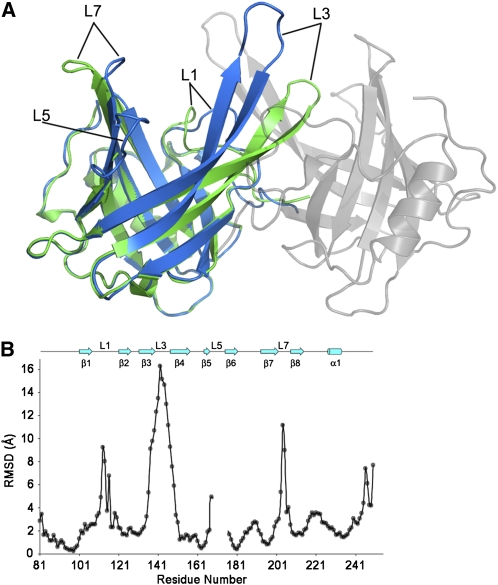
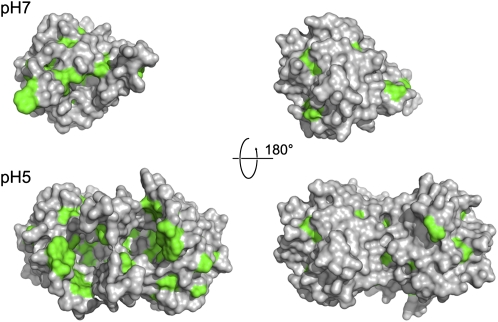
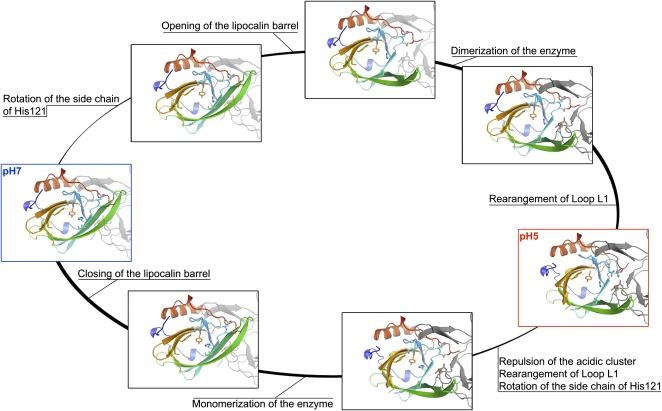
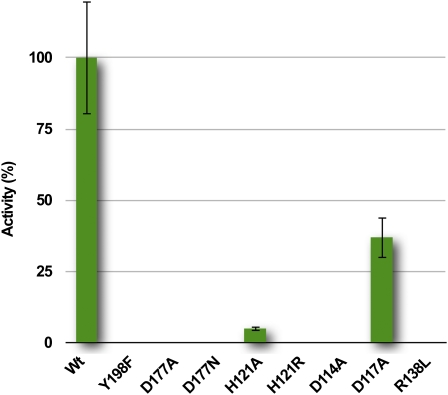
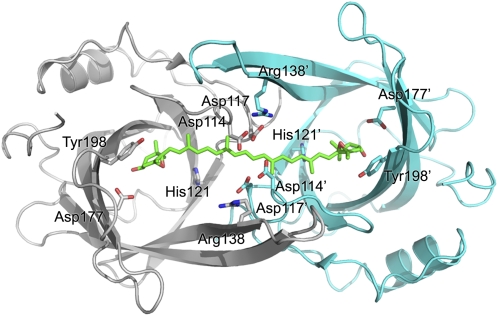
Plants adjust their photosynthetic activity to changing light conditions. A central regulation of photosynthesis depends on the xanthophyll cycle, in which the carotenoid violaxanthin is converted into zeaxanthin in strong light, thus activating the dissipation of the excess absorbed energy as heat and the scavenging of reactive oxygen species. Violaxanthin deepoxidase (VDE), the enzyme responsible for zeaxanthin synthesis, is activated by the acidification of the thylakoid lumen when photosynthetic electron transport exceeds the capacity of assimilatory reactions: at neutral pH, VDE is a soluble and inactive enzyme, whereas at acidic pH, it attaches to the thylakoid membrane where it binds its violaxanthin substrate. VDE also uses ascorbate as a cosubstrate with a pH-dependent Km that may reflect a preference for ascorbic acid. We determined the structures of the central lipocalin domain of VDE (VDEcd) at acidic and neutral pH. At neutral pH, VDEcd is monomeric with its active site occluded within a lipocalin barrel. Upon acidification, the barrel opens up and the enzyme appears as a dimer. A channel linking the two active sites of the dimer can harbor the entire carotenoid substrate and thus may permit the parallel deepoxidation of the two violaxanthin beta-ionone rings, making VDE an elegant example of the adaptation of an asymmetric enzyme to its symmetric substrate.
Figures
References
-
- Ahn, T.K., Avenson, T.J., Ballottari, M., Cheng, Y.C., Niyogi, K.K., Bassi, R., and Fleming, G.R. (2008). Architecture of a charge-transfer state regulating light harvesting in a plant antenna protein. Science 320 794–797. - PubMed
-
- Amarie, S., Laura, W., Tiago, B., Werner, K., Andreas, D., and Josef, W. (2009). Properties of zeaxanthin and its radical cation bound to the minor light-harvesting complexes CP24, CP26 and CP29. Biochimic. Biophys. Acta 1787 747–752. - PubMed
-
- Barber, J., and Andersson, B. (1992). Too much of a good thing: Light can be bad for photosynthesis. Trends Biochem. Sci. 17 61–66. - PubMed
-
- Bratt, C., Arvidsson, P., Carlsson, M., and Akerlund, H. (1995). Regulation of violaxanthin de-epoxidase activity by pH and ascorbate. Photosynth. Res. 45 169–175. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
