

Learning substrates in the primate prefrontal cortex and striatum: sustained activity related to successful actions
- PMID: 19640482
- PMCID: PMC2874751
- DOI: 10.1016/j.neuron.2009.06.019
Learning substrates in the primate prefrontal cortex and striatum: sustained activity related to successful actions
Abstract
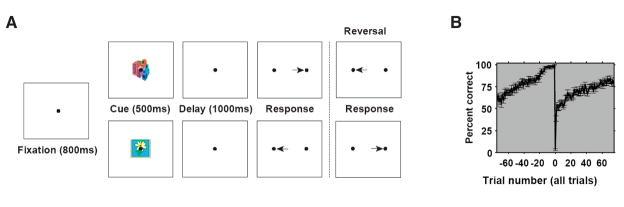
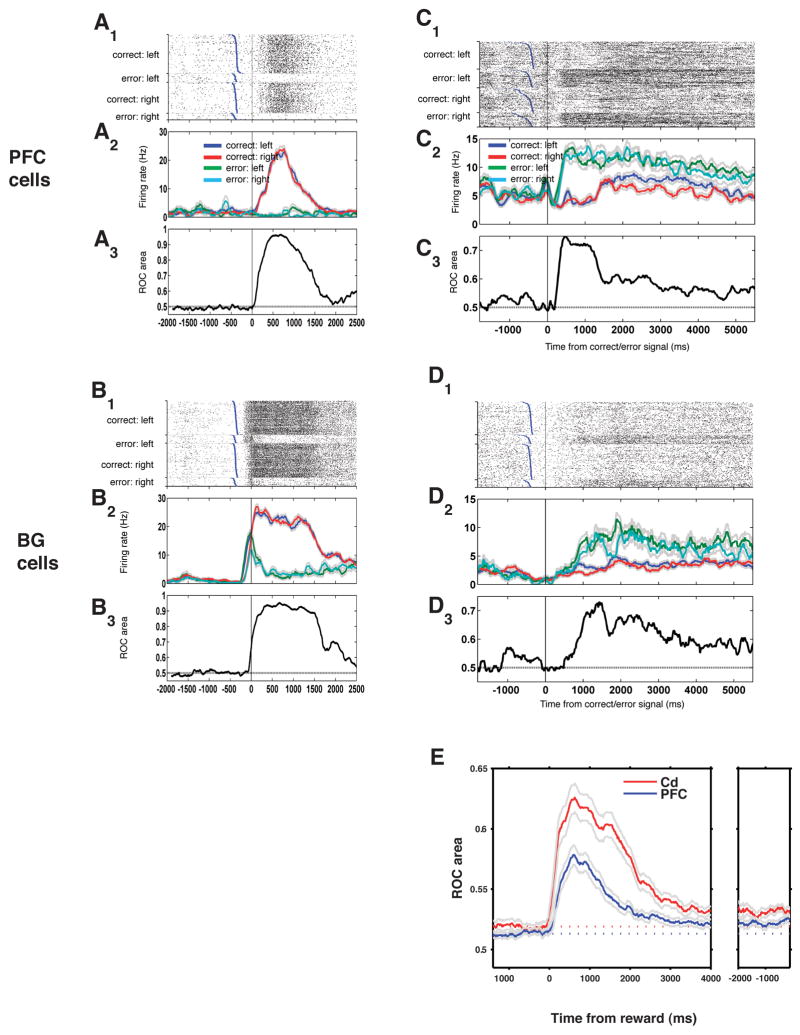
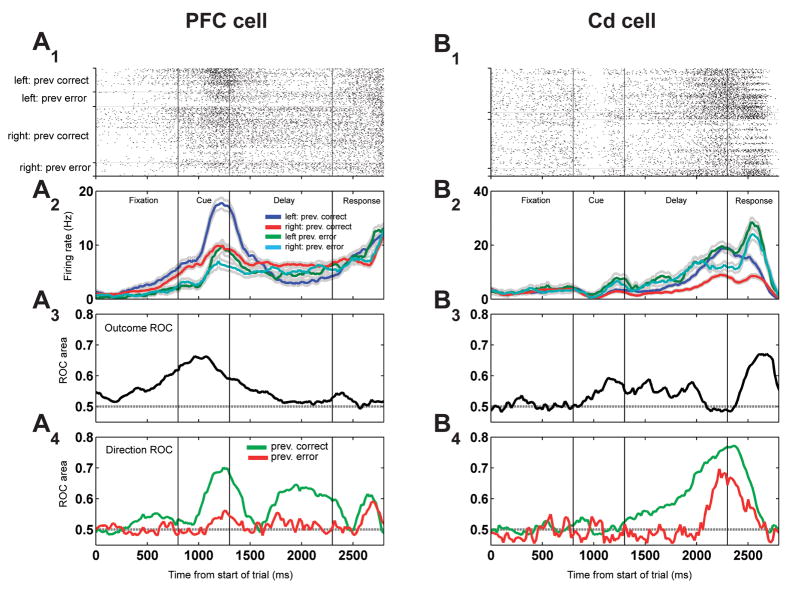
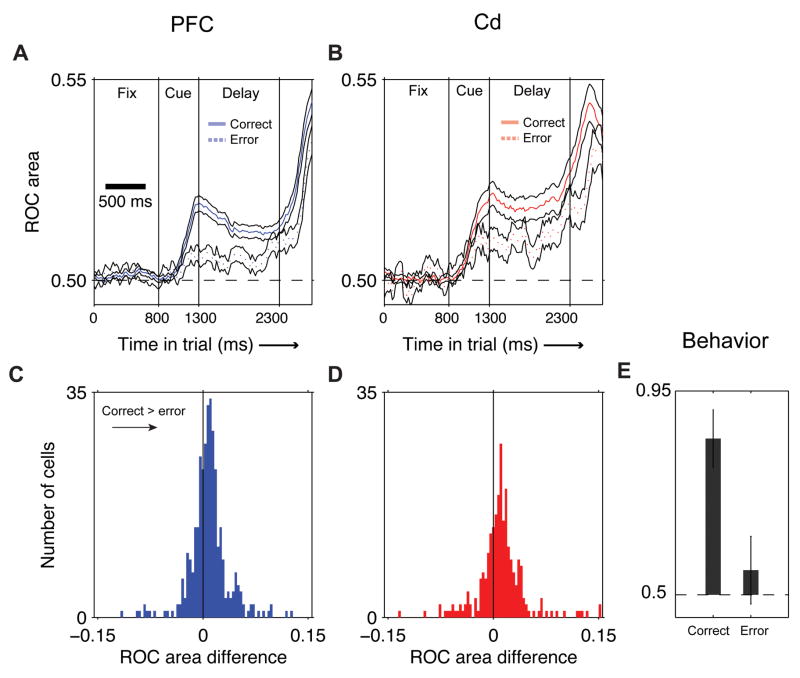
Learning from experience requires knowing whether a past action resulted in a desired outcome. The prefrontal cortex and basal ganglia are thought to play key roles in such learning of arbitrary stimulus-response associations. Previous studies have found neural activity in these areas, similar to dopaminergic neurons' signals, that transiently reflect whether a response is correct or incorrect. However, it is unclear how this transient activity, which fades in under a second, influences actions that occur much later. Here, we report that single neurons in both areas show sustained, persistent outcome-related responses. Moreover, single behavioral outcomes influence future neural activity and behavior: behavioral responses are more often correct and single neurons more accurately discriminate between the possible responses when the previous response was correct. These long-lasting signals about trial outcome provide a way to link one action to the next and may allow reward signals to be combined over time to implement successful learning.
Figures
Comment in
-
Past performance is indicative of future returns.Neuron. 2009 Jul 30;63(2):146-8. doi: 10.1016/j.neuron.2009.07.007. Neuron. 2009. PMID: 19640473
References
-
- Alexander GE, Crutcher MD, DeLong MR. Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, “prefrontal” and “limbic” functions. Prog Brain Res. 1990;85:119–146. - PubMed
-
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–381. - PubMed
-
- Anden NE, Hfuxe K, Hamberger B, Hokfelt T. A quantitative study on the nigro-neostriatal dopamine neuron system in the rat. Acta Physiol Scand. 1966;67:306–312. - PubMed
-
- Apicella P, Ljungberg T, Scarnati E, Schultz W. Responses to reward in monkey dorsal and ventral striatum. Exp Brain Res. 1991;85:491–500. - PubMed
-
- Asaad WF, Rainer G, Miller EK. Neural activity in the primate prefrontal cortex during associative learning. Neuron. 1998;21:1399–1407. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
