

Neuronal 3',3,5-triiodothyronine (T3) uptake and behavioral phenotype of mice deficient in Mct8, the neuronal T3 transporter mutated in Allan-Herndon-Dudley syndrome
- PMID: 19641107
- PMCID: PMC6666526
- DOI: 10.1523/JNEUROSCI.6055-08.2009
Neuronal 3',3,5-triiodothyronine (T3) uptake and behavioral phenotype of mice deficient in Mct8, the neuronal T3 transporter mutated in Allan-Herndon-Dudley syndrome
Abstract
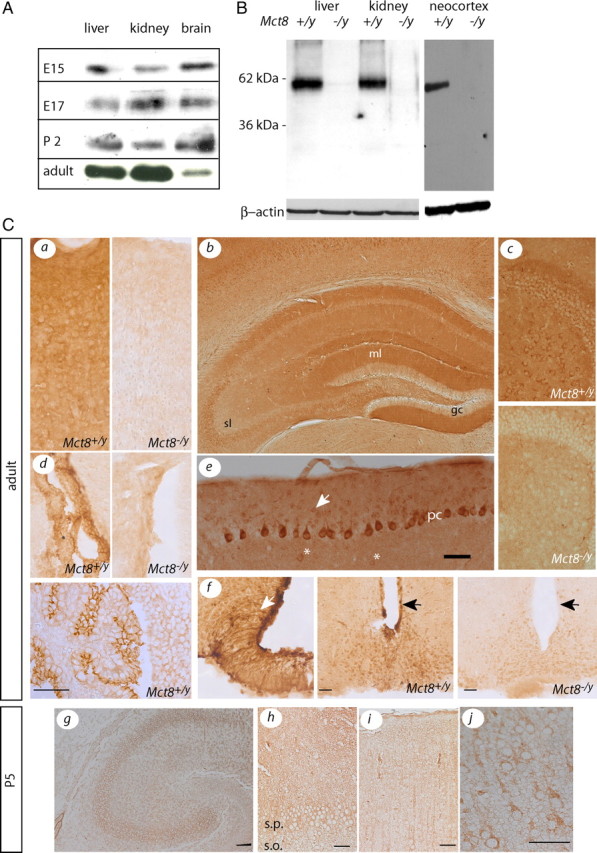
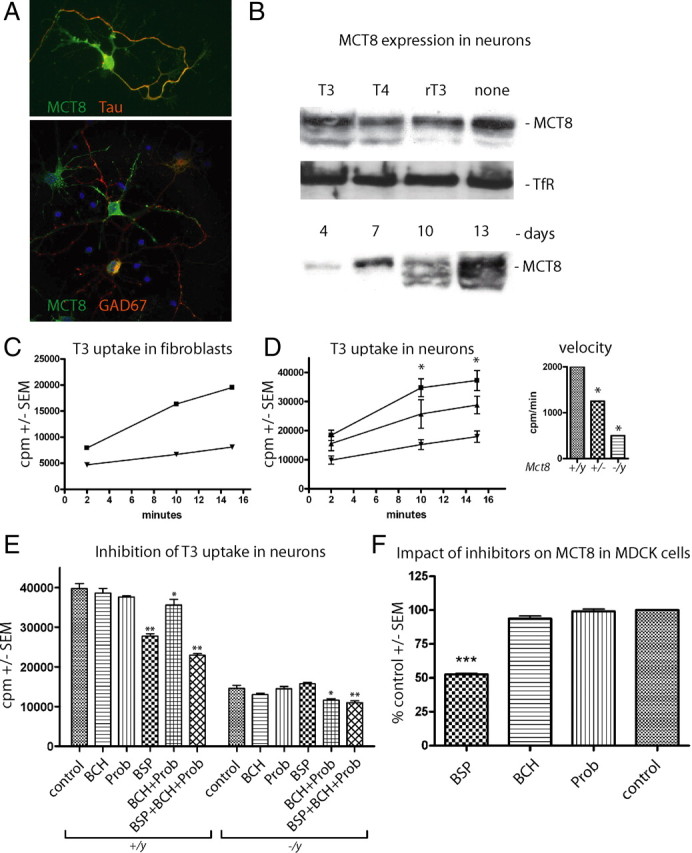
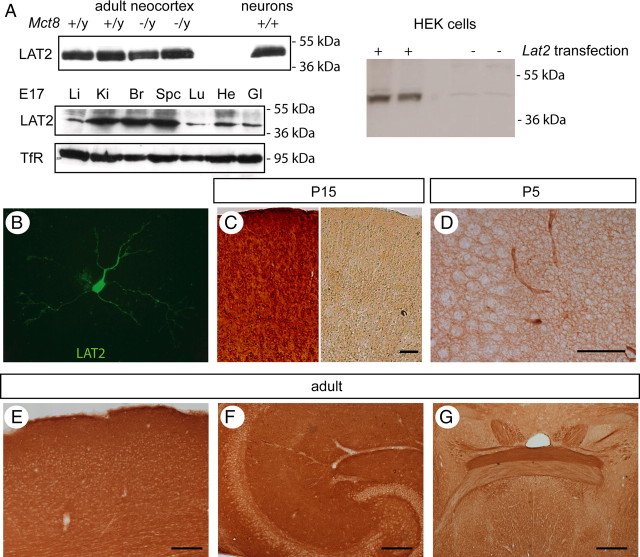
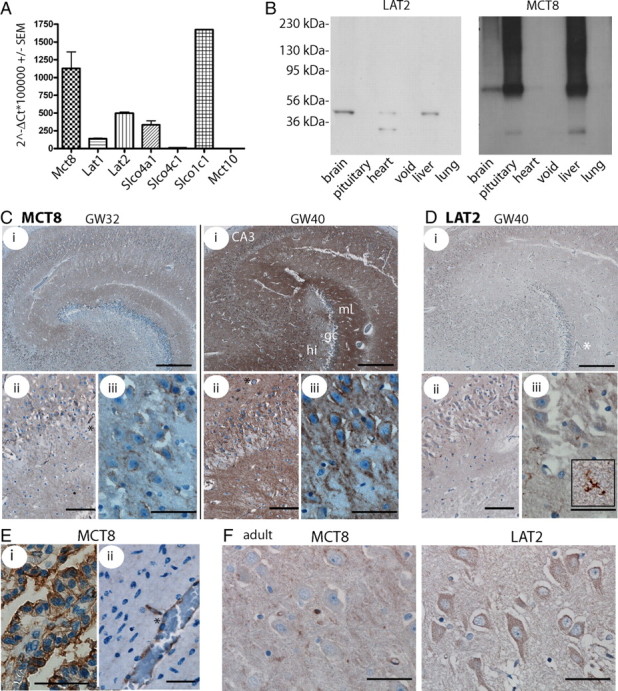
Thyroid hormone transport into cells requires plasma membrane transport proteins. Mutations in one of these, monocarboxylate transporter 8 (MCT8), have been identified as underlying cause for the Allan-Herndon-Dudley syndrome, an X-linked mental retardation in which the patients also present with abnormally high 3',3,5-triiodothyronine (T(3)) plasma levels. Mice deficient in Mct8 replicate the thyroid hormone abnormalities observed in the human condition. However, no neurological deficits have been described in mice lacking Mct8. Therefore, we subjected Mct8-deficient mice to a comprehensive immunohistochemical, neurological, and behavioral screen. Several behavioral abnormalities were found in the mutants. Interestingly, some of these behavioral changes are compatible with hypothyroidism, whereas others rather indicate hyperthyroidism. We thus hypothesized that neurons exclusively dependent on Mct8 are in a hypothyroid state, whereas neurons expressing other T(3) transporters become hyperthyroid, if they are exposed directly to the high plasma T(3). The majority of T(3) uptake in primary cortical neurons is mediated by Mct8, but pharmacological inhibition suggested functional expression of additional T(3) transporter classes. mRNAs encoding six T(3) transporters, including L-type amino acid transporters (LATs), were coexpressed with Mct8 in isolated neurons. We then demonstrated Lat2 expression in cultured neurons and throughout murine brain development. In contrast, LAT2 is expressed in microglia in the developing human brain during gestation, but not in neurons. We suggest that lack of functional complementation by alternative thyroid hormone transporters in developing human neurons precipitates the devastating neurodevelopmental phenotype in MCT8-deficient patients, whereas Mct8-deficient mouse neurons are functionally complemented by other transporters, for possibly Lat2.
Figures


References
-
- Ausó E, Lavado-Autric R, Cuevas E, Del Rey FE, Morreale De Escobar G, Berbel P. A moderate and transient deficiency of maternal thyroid function at the beginning of fetal neocorticogenesis alters neuronal migration. Endocrinology. 2004;145:4037–4047. - PubMed
-
- Bernal J. The significance of thyroid hormone transporters in the brain. Endocrinology. 2005a;146:1698–1700. - PubMed
-
- Bernal J. Thyroid hormones and brain development. Vitam Horm. 2005b;71:95–122. - PubMed
-
- Bernal J. Thyroid hormone receptors in brain development and function. Nat Clin Pract Endocrinol Metab. 2007;3:249–259. - PubMed
-
- Biebermann H, Ambrugger P, Tarnow P, von Moers A, Schweizer U, Grueters A. Extended clinical phenotype, endocrine investigations and functional studies of a loss-of-function mutation A150V in the thyroid hormone specific transporter MCT8. Eur J Endocrinol. 2005;153:359–366. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous