

doi: 10.1038/emboj.2009.213.
Epub 2009 Jul 30.
36 degrees step size of proton-driven c-ring rotation in FoF1-ATP synthase
Affiliations
- PMID: 19644443
- PMCID: PMC2750017
- DOI: 10.1038/emboj.2009.213
Item in Clipboard
36 degrees step size of proton-driven c-ring rotation in FoF1-ATP synthase
EMBO J.
.
Abstract
Synthesis of adenosine triphosphate ATP, the 'biological energy currency', is accomplished by F(o)F(1)-ATP synthase. In the plasma membrane of Escherichia coli, proton-driven rotation of a ring of 10 c subunits in the F(o) motor powers catalysis in the F(1) motor. Although F(1) uses 120 degrees stepping during ATP synthesis, models of F(o) predict either an incremental rotation of c subunits in 36 degrees steps or larger step sizes comprising several fast substeps. Using single-molecule fluorescence resonance energy transfer, we provide the first experimental determination of a 36 degrees sequential stepping mode of the c-ring during ATP synthesis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

Single-molecule FRET approach to detect the 36° step size of the rotary c subunits in FoF1-ATP synthase during ATP synthesis. (A) Model of FRET-labelled E. coli FoF1-ATP synthase with EGFP (green; fused to the C terminus of subunit a, orange) and Alexa568 (red, at residue E2C) at one of the c subunits (blue). Rotation of c results in stepwise distance changes to EGFP. (B) The FRET-labelled E. coli FoF1-ATP synthase is reconstituted and diffuses freely through the dual laser foci of the duty cycle-optimized alternating laser excitation scheme. (C) Photon burst of a single FRET-labelled FoF1-ATP synthase. In the lower panel, fluorescence intensities of FRET donor EGFP (green trace) and acceptor Alexa568 (red trace) are shown excited with 488 nm. In the upper panel, the fluorescence intensity of the directly excited Alexa568 (pulsed interleaved excitation with 561 nm, see text) of the same FoF1-ATP synthase is shown (black trace) confirming the existence of both markers on the enzyme.
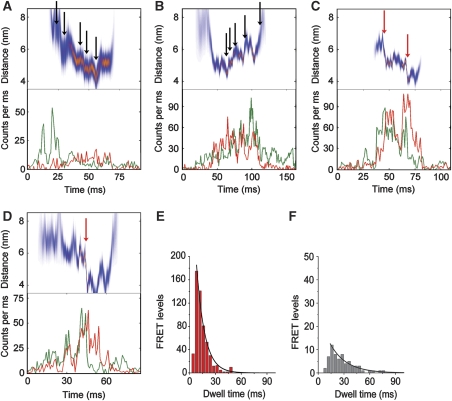
(A–D) Photon bursts of single FRET-labelled FoF1-ATP synthases during ATP synthesis. Lower panels show fluorescence time trajectories for EGFP (green trace) and Alexa568 (red trace), upper panels show the corresponding intramolecular FRET distances (most-likely distance in red and deviations as blue band). Black arrows mark small steps, red arrows large steps. (E) Dwell time distributions of FRET levels in FoF1-ATP synthases during ATP synthesis (red bars), and (F), with 20 μm aurovertin B (gray bars, monoexponential fits in black). Adding an additional rising component apparently improved the fitting at shorter dwells. This is due to the fact that the lower limit to determining dwell times is 2 ms, and thus the histograms lack those data points.

Model for the geometrical constraints for the FRET measurements within a single FoF1-ATP synthase and FRET transition density plot. (A) Model for FRET distances between the EGFP chromophore (green dot) and three of the 10 stopping positions of Alexa568 (red dots) during c-ring rotation. Distance changes for a 36° step correspond to arrows blue and cyan; a 144° step corresponds to arrows blue and black. (B) Five zones for the 10 c positions as defined from symmetry reasons. 36° stepping of c results in changing between adjacent zones, and, correspondingly, incremental FRET distance changes. (C) Larger step sizes of 108° require switching between non-neighbouring zones. (D) Transitions between FRET distances according to 36° steps and geometries as shown in panel B. (E) Transitions between FRET distances according to 120° steps. (F) FRET transition density plot for proton-driven c subunit rotation with constraint curves for 36° steps in white and for 120° steps in black.

Monte Carlo simulations of FRET distance changes during stepwise c-ring rotation and comparison with experimental data during ATP synthesis. (A) Experimental FRET distance changes. Simulations with 36° (B), 72° (C), 108° (D), 144° (E), 120° (F), 40° plus 80° (G); the weighted sum of 36°, 72°, 108° and 144° step sizes (H) reproduced the FRET distance difference plot and the experimental FRET transition density plot (see Figure 3F) fairly well. I, FRET transition density plot from the Monte Carlo simulation shown in (H).
Similar articles
-
Biased Brownian stepping rotation of FoF1-ATP synthase driven by proton motive force.Nat Commun. 2013;4:1631. doi: 10.1038/ncomms2631. Nat Commun. 2013. PMID: 23535652
-
Structural model of the transmembrane Fo rotary sector of H+-transporting ATP synthase derived by solution NMR and intersubunit cross-linking in situ.Biochim Biophys Acta. 2002 Oct 11;1565(2):232-45. doi: 10.1016/s0005-2736(02)00572-2. Biochim Biophys Acta. 2002. PMID: 12409198 Review.
-
Operation mechanism of F(o) F(1)-adenosine triphosphate synthase revealed by its structure and dynamics.IUBMB Life. 2013 Mar;65(3):238-46. doi: 10.1002/iub.1120. Epub 2013 Jan 22. IUBMB Life. 2013. PMID: 23341301 Review.
-
The regulatory subunit ε in Escherichia coli FOF1-ATP synthase.Biochim Biophys Acta Bioenerg. 2018 Sep;1859(9):775-788. doi: 10.1016/j.bbabio.2018.06.013. Epub 2018 Jun 20. Biochim Biophys Acta Bioenerg. 2018. PMID: 29932911 Free PMC article. Review.
-
Cooperation among c-subunits of FoF1-ATP synthase in rotation-coupled proton translocation.Elife. 2022 Feb 2;11:e69096. doi: 10.7554/eLife.69096. Elife. 2022. PMID: 35107420 Free PMC article.
Cited by
-
Biased Brownian stepping rotation of FoF1-ATP synthase driven by proton motive force.Nat Commun. 2013;4:1631. doi: 10.1038/ncomms2631. Nat Commun. 2013. PMID: 23535652
-
FO-F1 coupling and symmetry mismatch in ATP synthase resolved in every FO rotation step.Biophys J. 2023 Jul 25;122(14):2898-2909. doi: 10.1016/j.bpj.2022.09.034. Epub 2022 Sep 28. Biophys J. 2023. PMID: 36171725 Free PMC article.
-
ATP synthase: from single molecule to human bioenergetics.Proc Jpn Acad Ser B Phys Biol Sci. 2010;86(7):667-93. doi: 10.2183/pjab.86.667. Proc Jpn Acad Ser B Phys Biol Sci. 2010. PMID: 20689227 Free PMC article.
-
The regulatory switch of F1-ATPase studied by single-molecule FRET in the ABEL Trap.Proc SPIE Int Soc Opt Eng. 2014 Apr 1;8950:89500H. doi: 10.1117/12.2042688. Proc SPIE Int Soc Opt Eng. 2014. PMID: 25309100 Free PMC article.
-
High-resolution single-molecule characterization of the enzymatic states in Escherichia coli F1-ATPase.Philos Trans R Soc Lond B Biol Sci. 2012 Dec 24;368(1611):20120023. doi: 10.1098/rstb.2012.0023. Print 2013 Feb 5. Philos Trans R Soc Lond B Biol Sci. 2012. PMID: 23267177 Free PMC article.
References
-
- Aggeler R, Ogilvie I, Capaldi RA (1997) Rotation of a gamma-epsilon subunit domain in the Escherichia coli F1F0-ATP synthase complex. The gamma-epsilon subunits are essentially randomly distributed relative to the alpha3beta3delta domain in the intact complex. J Biol Chem 272: 19621–19624 - PubMed
-
- Antonik M, Felekyan S, Gaiduk A, Seidel CA (2006) Separating structural heterogeneities from stochastic variations in fluorescence resonance energy transfer distributions via photon distribution analysis. J Phys Chem B 110: 6970–6978 - PubMed
-
- Borsch M, Diez M, Zimmermann B, Reuter R, Graber P (2002) Stepwise rotation of the gamma-subunit of EF(0)F(1)-ATP synthase observed by intramolecular single-molecule fluorescence resonance energy transfer. FEBS Lett 527: 147–152 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
