

Identification of a perinuclear positioning element in human subtelomeres that requires A-type lamins and CTCF
- PMID: 19644448
- PMCID: PMC2735173
- DOI: 10.1038/emboj.2009.201
Identification of a perinuclear positioning element in human subtelomeres that requires A-type lamins and CTCF
Abstract
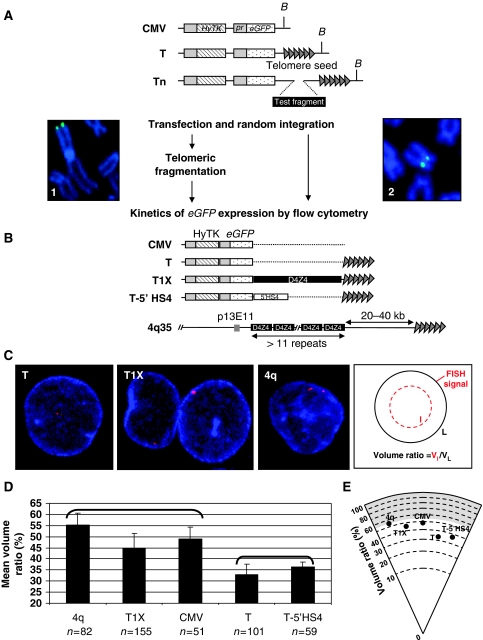
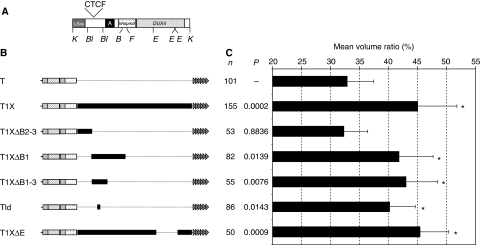
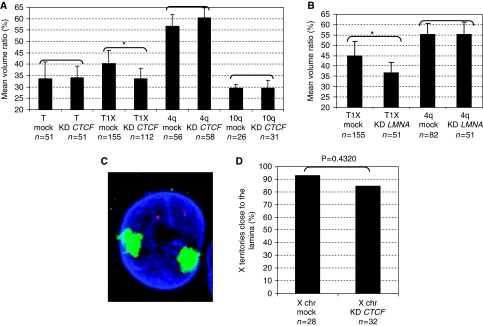
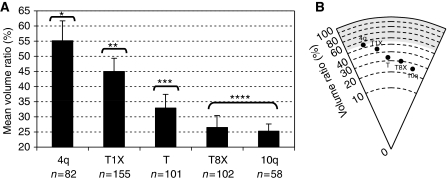
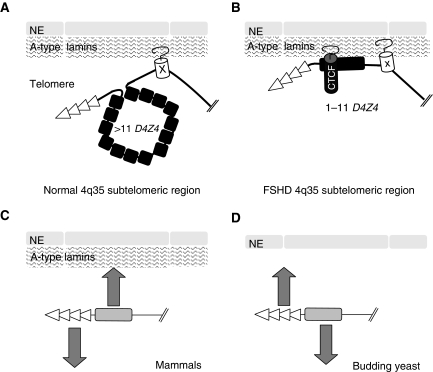
The localization of genes within the nuclear space is of paramount importance for proper genome functions. However, very little is known on the cis-acting elements determining subnuclear positioning of chromosome segments. We show here that the D4Z4 human subtelomeric repeat localizes a telomere at the nuclear periphery. This perinuclear activity lies within an 80 bp sequence included within a region known to interact with CTCF and A-type Lamins. We further show that a reduced level of either CTCF or A-type Lamins suppresses the perinuclear activities of D4Z4 and that an array of multimerized D4Z4 sequence, which has lost its ability to bind CTCF and A-type Lamins, is not localized at the periphery. Overall, these findings reveal the existence of an 80 bp D4Z4 sequence that is sufficient to position an adjacent telomere to the nuclear periphery in a CTCF and A-type lamins-dependent manner. Strikingly, this sequence includes a 30 bp GA-rich motif, which binds CTCF and is present at several locations in the human genome.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Andrulis ED, Neiman AM, Zappulla DC, Sternglanz R (1998) Perinuclear localization of chromatin facilitates transcriptional silencing. Nature 394: 592–595 - PubMed
-
- Bakay M, Wang Z, Melcon G, Schiltz L, Xuan J, Zhao P, Sartorelli V, Seo J, Pegoraro E, Angelini C, Shneiderman B, Escolar D, Chen YW, Winokur ST, Pachman LM, Fan C, Mandler R, Nevo Y, Gordon E, Zhu Y et al. (2006) Nuclear envelope dystrophies show a transcriptional fingerprint suggesting disruption of Rb-MyoD pathways in muscle regeneration. Brain 129 (Pt 4): 996–1013 - PubMed
-
- Brown KE, Guest SS, Smale ST, Hahm K, Merkenschlager M, Fisher AG (1997) Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin. Cell 91: 845–854 - PubMed
-
- Chung JH, Whiteley M, Felsenfeld G (1993) A 5′ element of the chicken beta-globin domain serves as an insulator in human erythroid cells and protects against position effect in Drosophila. Cell 74: 505–514 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
