

Steering electrons on moving pathways
- PMID: 19645446
- PMCID: PMC2764794
- DOI: 10.1021/ar900123t
Steering electrons on moving pathways
Abstract
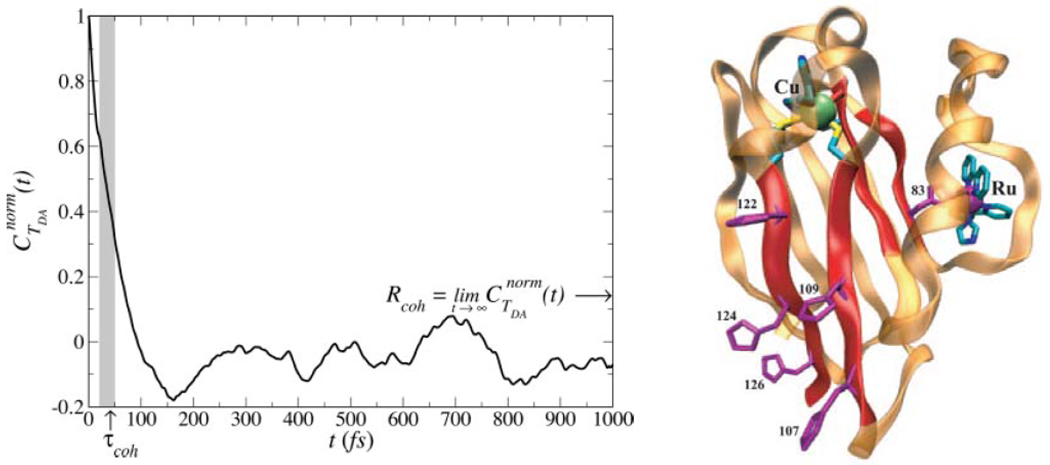
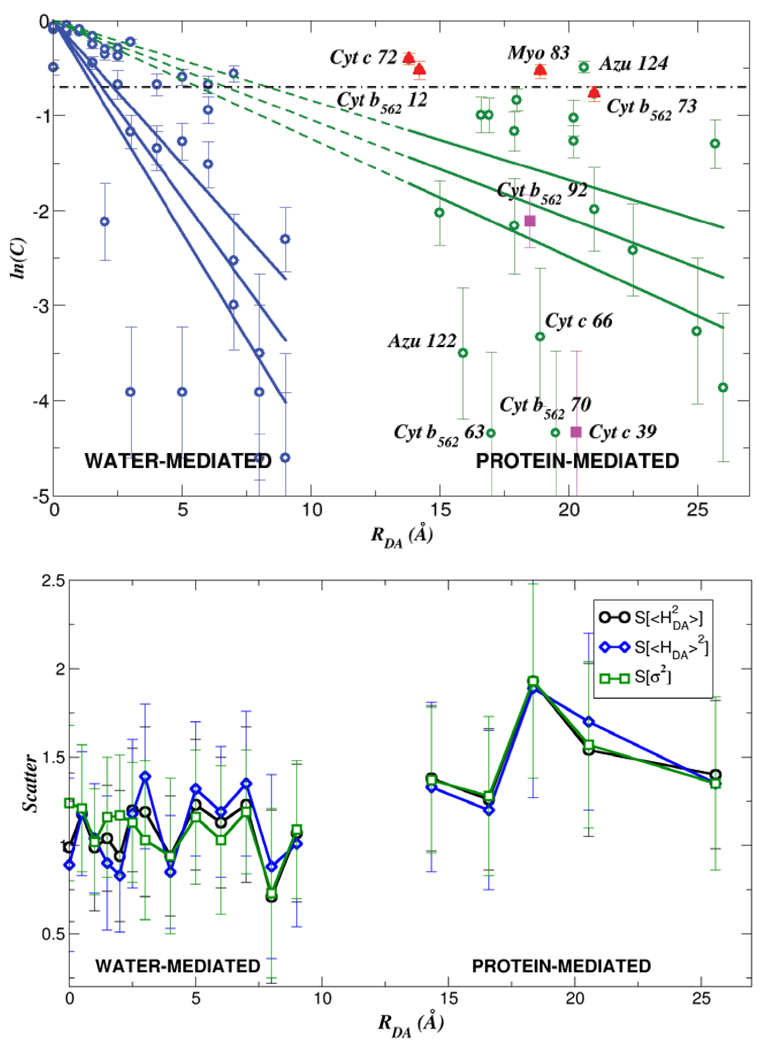
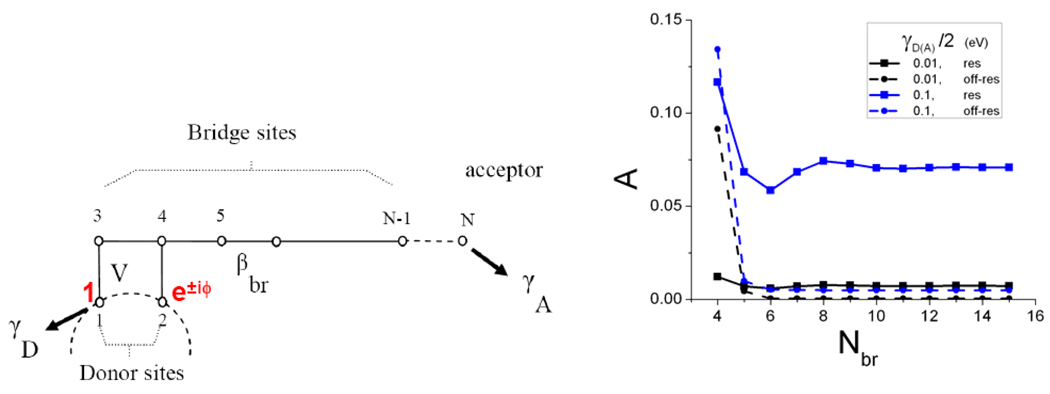
Electron transfer (ET) reactions provide a nexus among chemistry, biochemistry, and physics. These reactions underpin the "power plants" and "power grids" of bioenergetics, and they challenge us to understand how evolution manipulates structure to control ET kinetics. Ball-and-stick models for the machinery of electron transfer, however, fail to capture the rich electronic and nuclear dynamics of ET molecules: these static representations disguise, for example, the range of thermally accessible molecular conformations. The influence of structural fluctuations on electron-transfer kinetics is amplified by the exponential decay of electron tunneling probabilities with distance, as well as the delicate interference among coupling pathways. Fluctuations in the surrounding medium can also switch transport between coherent and incoherent ET mechanisms--and may gate ET so that its kinetics is limited by conformational interconversion times, rather than by the intrinsic ET time scale. Moreover, preparation of a charge-polarized donor state or of a donor state with linear or angular momentum can have profound dynamical and kinetic consequences. In this Account, we establish a vocabulary to describe how the conformational ensemble and the prepared donor state influence ET kinetics in macromolecules. This framework is helping to unravel the richness of functional biological ET pathways, which have evolved within fluctuating macromolecular structures. The conceptual framework for describing nonadiabatic ET seems disarmingly simple: compute the ensemble-averaged (mean-squared) donor-acceptor (DA) tunneling interaction, <H(DA)(2)>, and the Franck-Condon weighted density of states, rho(FC), to describe the rate, (2pi/variant Planck's over 2pi)<H(DA)(2)>rho(FC). Modern descriptions of the thermally averaged electronic coupling and of the Franck-Condon factor establish a useful predictive framework in biology, chemistry, and nanoscience. Describing the influence of geometric and energetic fluctuations on ET allows us to address a rich array of mechanistic and kinetic puzzles. How strongly is a protein's fold imprinted on the ET kinetics, and might thermal fluctuations "wash out" signatures of structure? What is the influence of thermal fluctuations on ET kinetics beyond averaging of the tunneling barrier structure? Do electronic coupling mechanisms change as donor and acceptor reposition in a protein, and what are the consequences for the ET kinetics? Do fluctuations access minority species that dominate tunneling? Can energy exchanges between the electron and bridge vibrations generate vibronic signatures that label some of the D-to-A pathways traversed by the electron, thus eliminating unmarked pathways that would otherwise contribute to the DA coupling (as in other "which way" or double-slit experiments)? Might medium fluctuations drive tunneling-hopping mechanistic transitions? How does the donor-state preparation, in particular, its polarization toward the acceptor and its momentum characteristics (which may introduce complex rather than pure real relationships among donor orbital amplitudes), influence the electronic dynamics? In this Account, we describe our recent studies that address puzzling questions of how conformational distributions, excited-state polarization, and electronic and nuclear dynamical effects influence ET in macromolecules. Indeed, conformational and dynamical effects arise in all transport regimes, including the tunneling, resonant transport, and hopping regimes. Importantly, these effects can induce switching among ET mechanisms.
Figures




References
-
- Bendall DS, editor. Protein Electron Transfer. Oxford: BIOS Scientific Publishers, Inc.; 1996.
-
- Skourtis SS, Lin J, Beratan DN. In: Modern Methods for theoretical Physical Chemistry of Biopolymers. Starikov EB, Lewis JP, Tanaka S, editors. Boston: Elsevier; 2006.
-
- Nitzan A. Electron transmission through molecules and molecular interfaces. Annu. Rev. Phys. Chem. 2001;52:681–750. - PubMed
-
- Bixon M, Jortner J, editors. Adv. Chem. Phys. 1999. pp. 106–107. Vol.
-
- Newton MD. In: Electron Transfer in Chemistry. Balzani V, editor. Vol. 1. New York: Wiley-VCH; 2001. pp. 3–63.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
