

Mechanosensing through cooperative interactions between myosin II and the actin crosslinker cortexillin I
- PMID: 19646871
- PMCID: PMC2763054
- DOI: 10.1016/j.cub.2009.07.018
Mechanosensing through cooperative interactions between myosin II and the actin crosslinker cortexillin I
Abstract
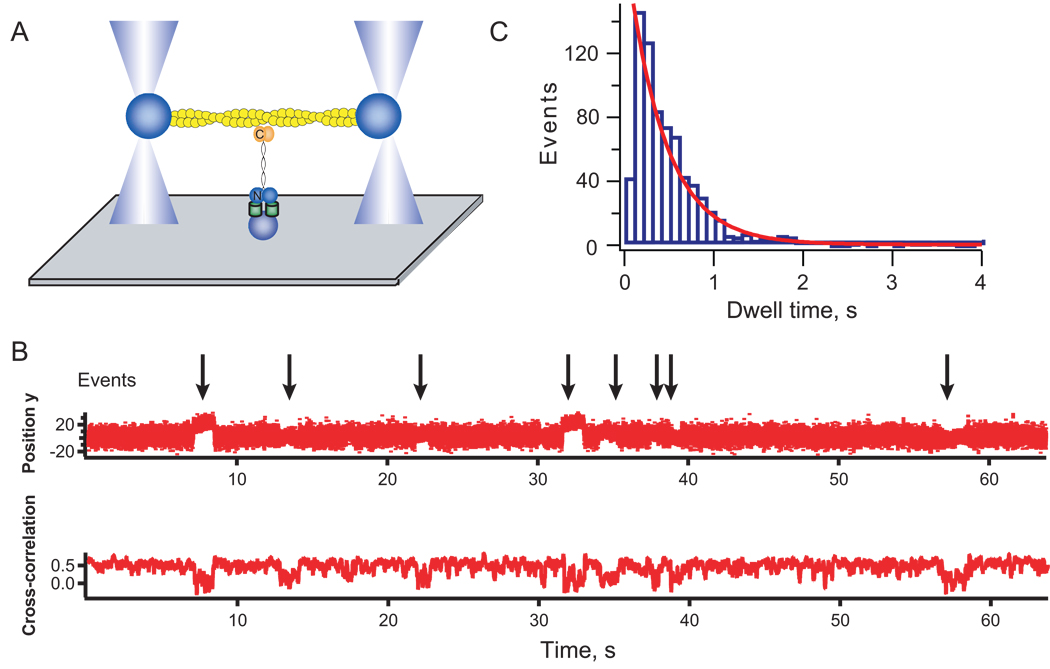
Background: Mechanosensing governs many processes from molecular to organismal levels, including during cytokinesis where it ensures successful and symmetrical cell division. Although many proteins are now known to be force sensitive, myosin motors with their ATPase activity and force-sensitive mechanical steps are well poised to facilitate cellular mechanosensing. For a myosin motor to experience tension, the actin filament must also be anchored.
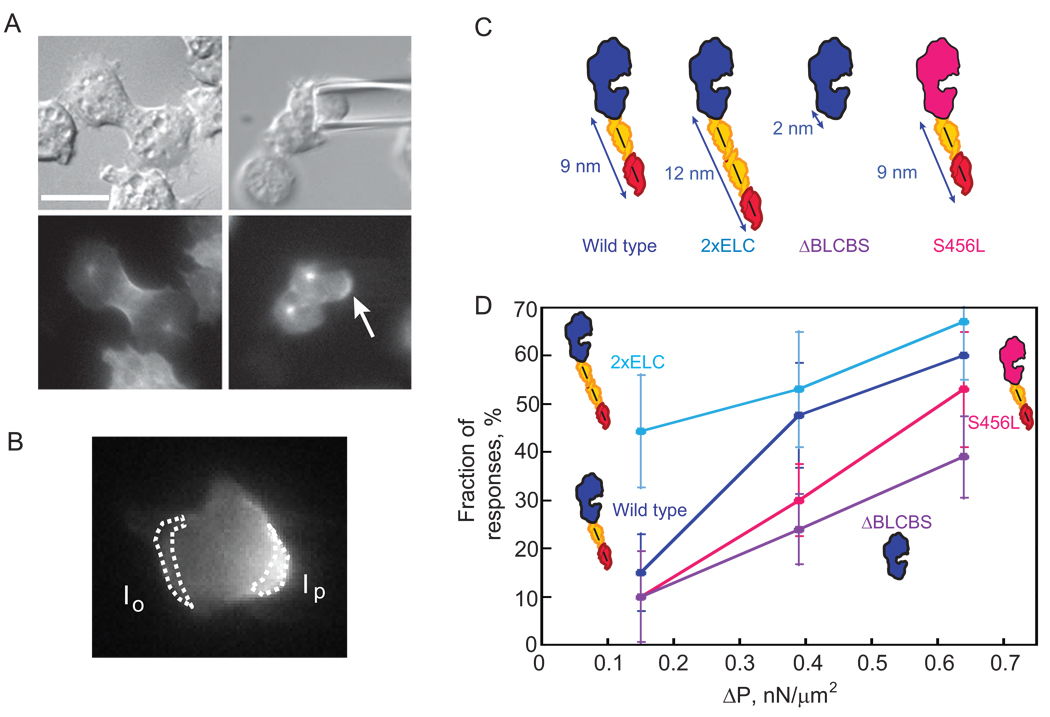
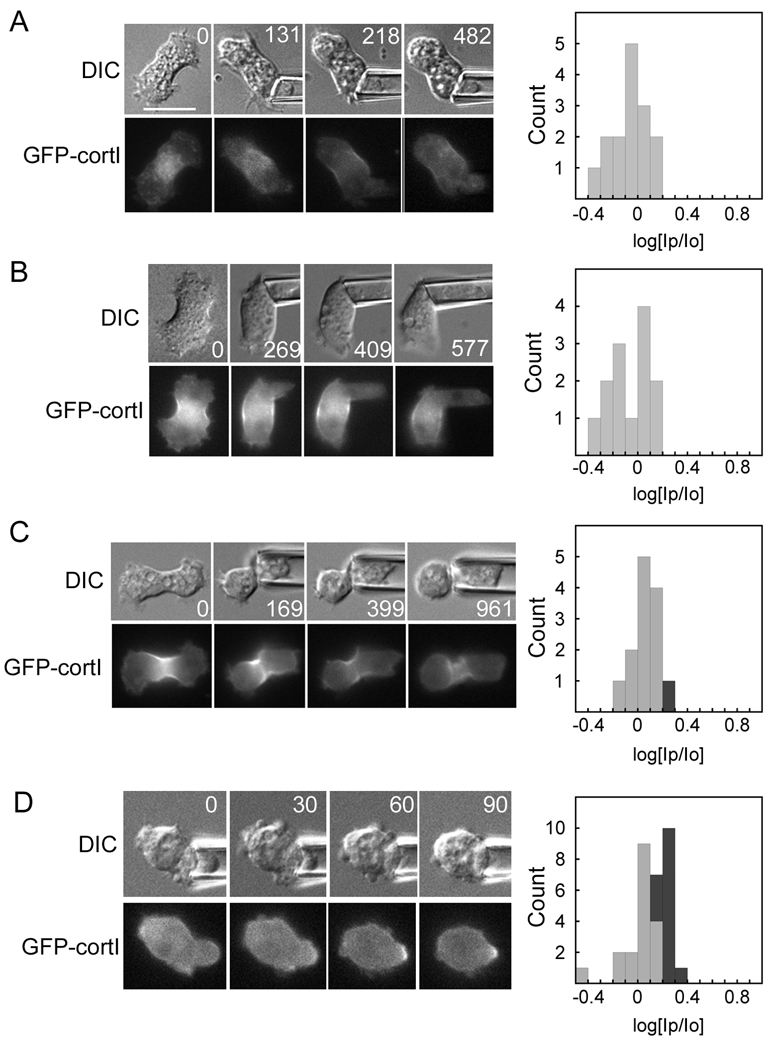
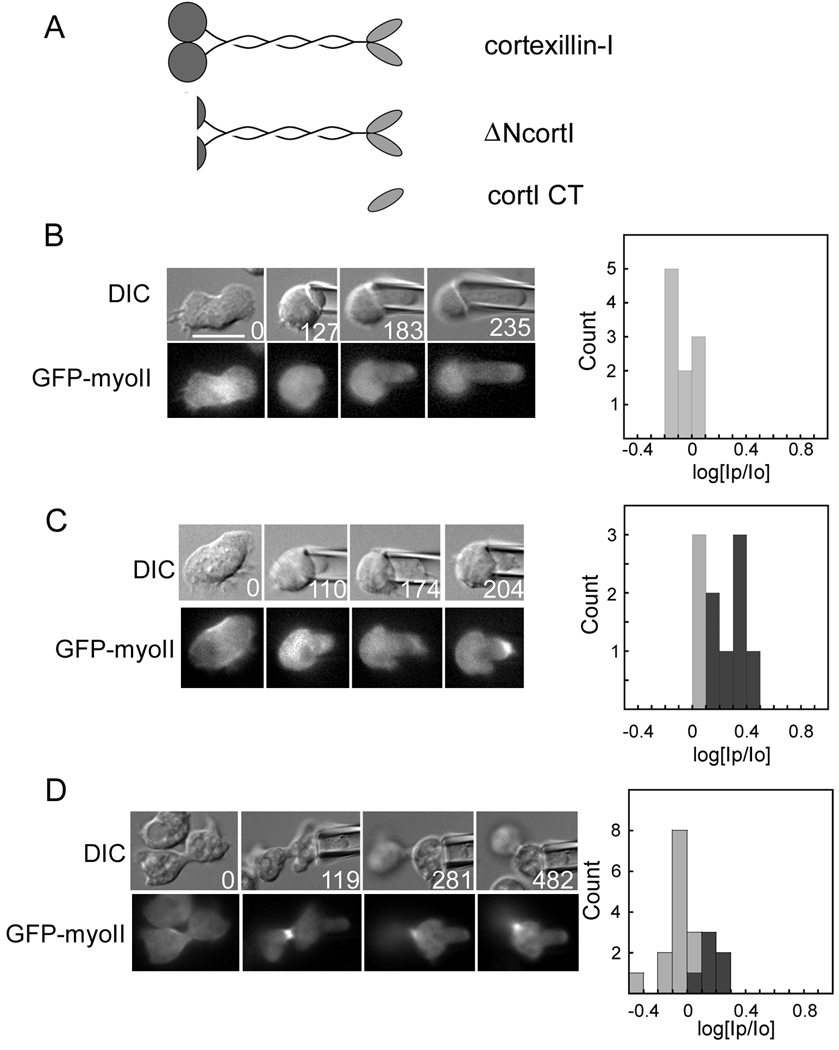
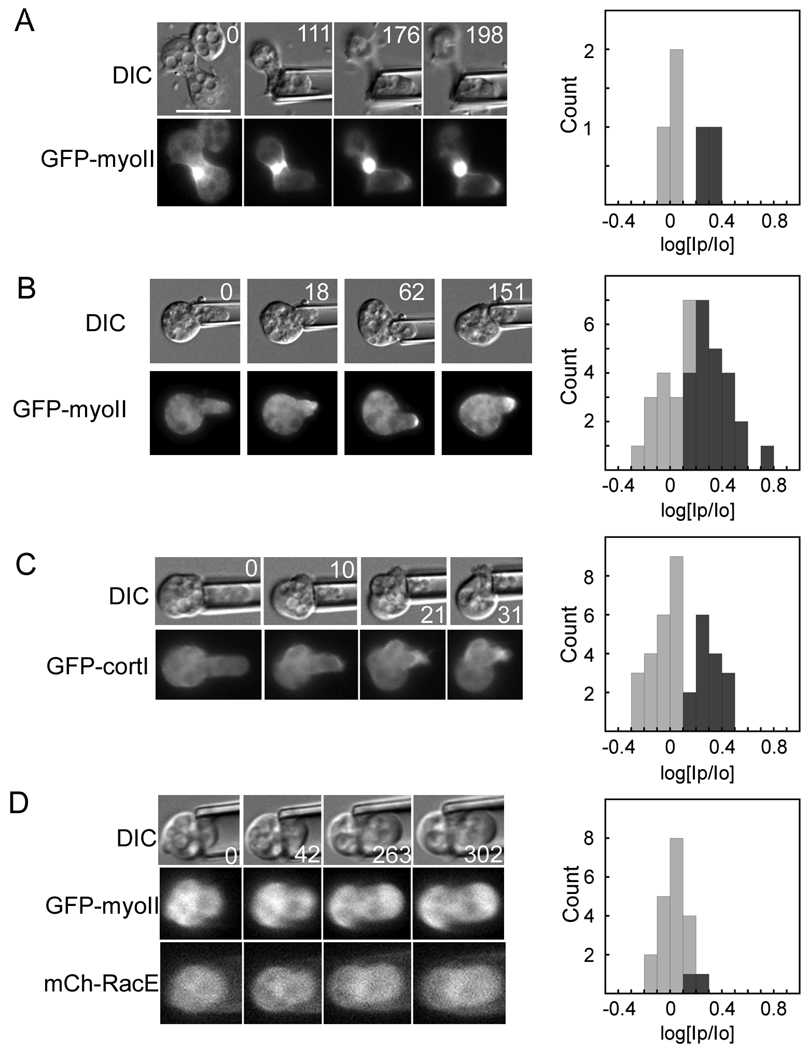
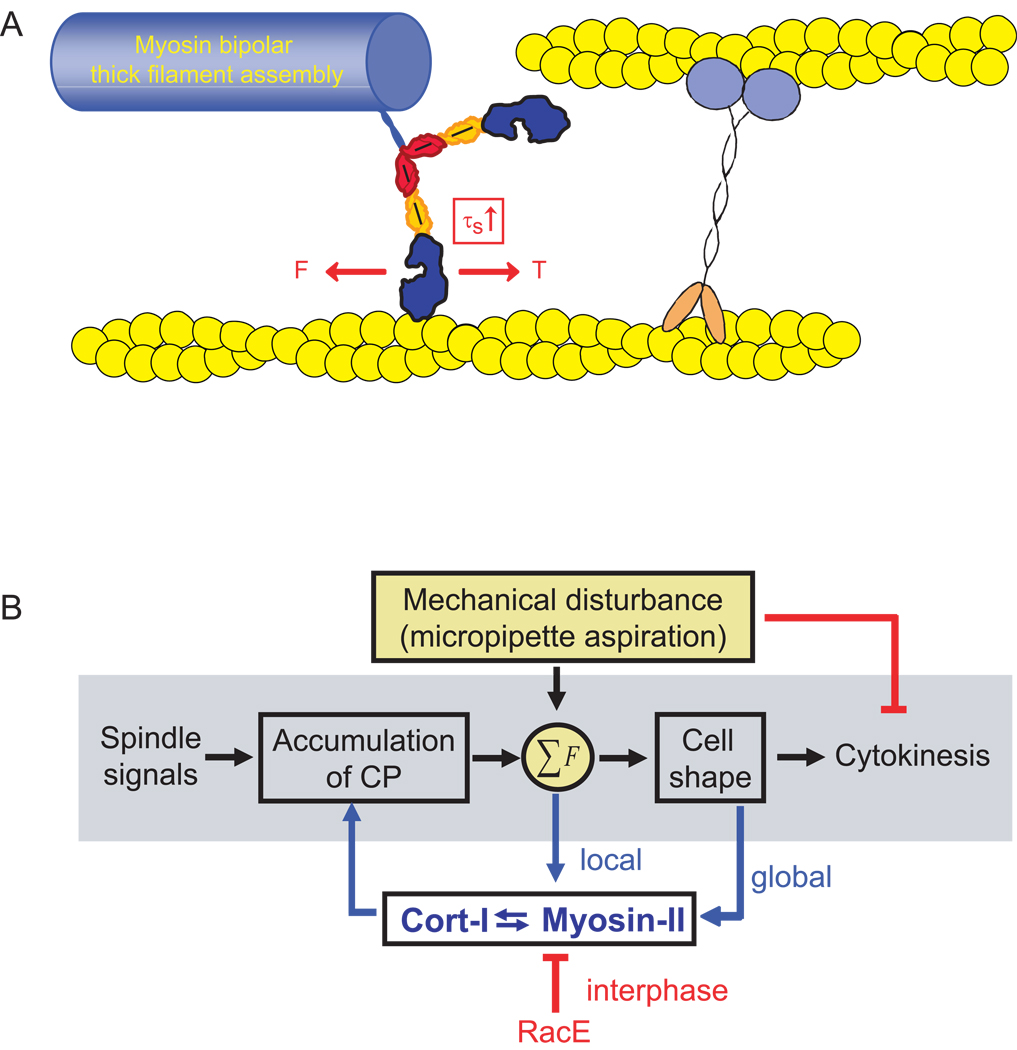
Results: Here, we find a cooperative relationship between myosin II and the actin crosslinker cortexillin I where both proteins are essential for cellular mechanosensory responses. Although many functions of cortexillin I and myosin II are dispensable for cytokinesis, all are required for full mechanosensing. Our analysis demonstrates that this mechanosensor has three critical elements: the myosin motor where the lever arm acts as a force amplifier, a force-sensitive bipolar thick-filament assembly, and a long-lived actin crosslinker, which anchors the actin filament so that the motor may experience tension. We also demonstrate that a Rac small GTPase inhibits this mechanosensory module during interphase, allowing the module to be primarily active during cytokinesis.
Conclusions: Overall, myosin II and cortexillin I define a cellular-scale mechanosensor that controls cell shape during cytokinesis. This system is exquisitely tuned through the enzymatic properties of the myosin motor, its lever arm length, and bipolar thick-filament assembly dynamics. The system also requires cortexillin I to stably anchor the actin filament so that the myosin motor can experience tension. Through this cross-talk, myosin II and cortexillin I define a cellular-scale mechanosensor that monitors and corrects shape defects, ensuring symmetrical cell division.
Figures
References
-
- Orr AW, Helmke BP, Blackman BR, Schwartz MA. Mechanisms of mechanotransduction. Dev. Cell. 2006;10:11–20. - PubMed
-
- Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126:677–689. - PubMed
-
- Martinac B. Mechanosensitive ion channels: molecules of mechanotransduction. J. Cell Sci. 2004;117:2449–2460. - PubMed
-
- Balaban NQ, Schwartz US, Riveline D, Goichberg P, Tzur G, Sabanay I, Mahalu D, Safran S, Bershadsky AD, Addadi L, Geiger B. Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 2001;3:466–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
