

Augmented potassium current is a shared phenotype for two genetic defects associated with familial atrial fibrillation
- PMID: 19646991
- PMCID: PMC2813326
- DOI: 10.1016/j.yjmcc.2009.07.020
Augmented potassium current is a shared phenotype for two genetic defects associated with familial atrial fibrillation
Abstract
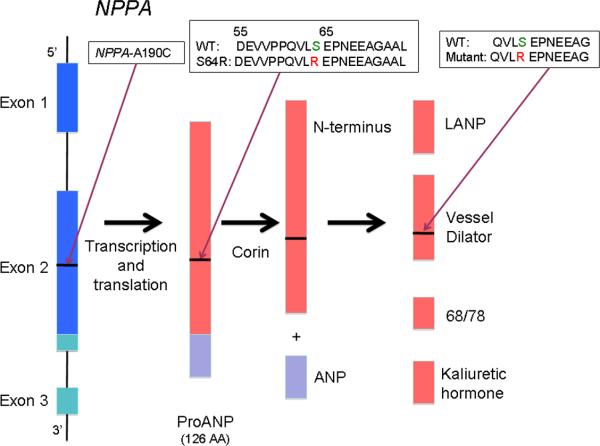
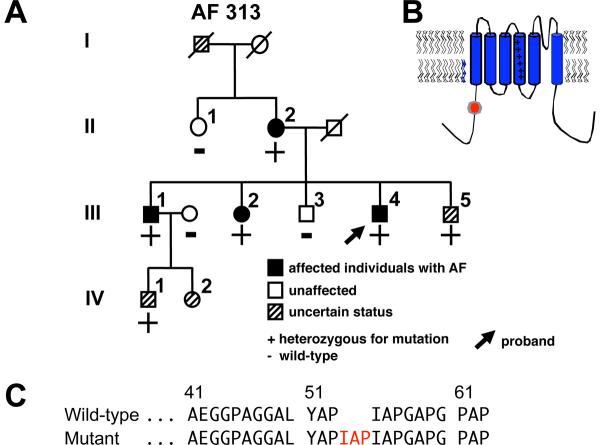
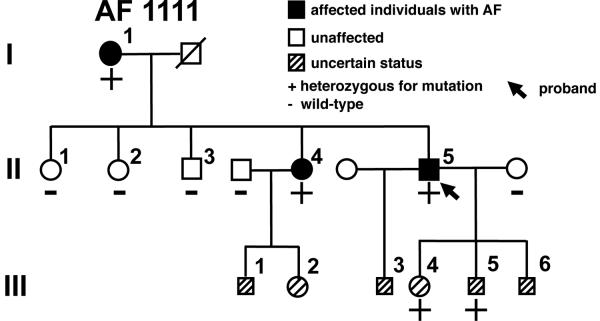
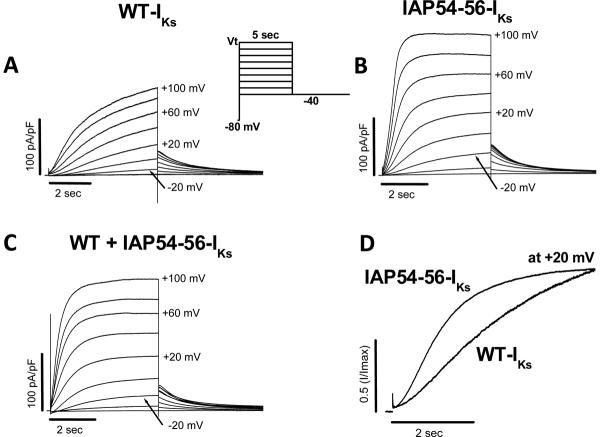
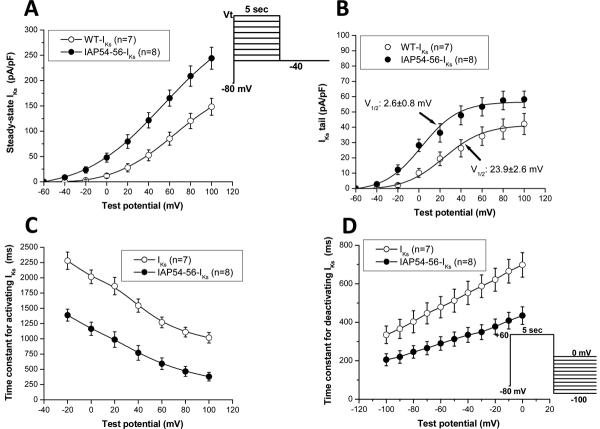
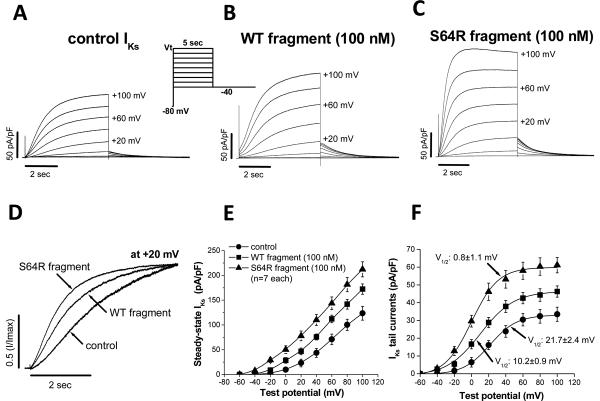
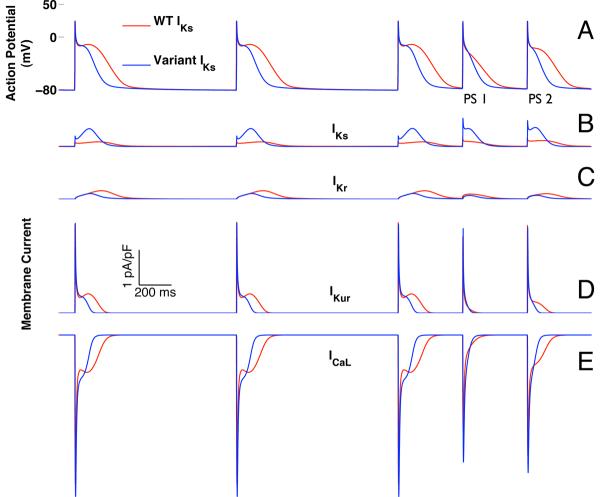
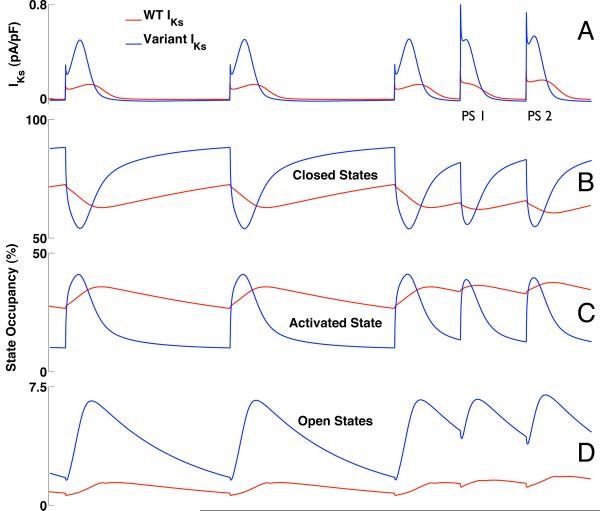
Mutations in multiple genes have been implicated in familial atrial fibrillation (AF), but the underlying mechanisms, and thus implications for therapy, remain ill-defined. Among 231 participants in the Vanderbilt AF Registry, we found a mutation in KCNQ1 (encoding the alpha-subunit of slow delayed rectifier potassium current [I(Ks)]) and separately a mutation in natriuretic peptide precursor A (NPPA) gene (encoding atrial natriuretic peptide, ANP), both segregating with early onset lone AF in different kindreds. The functional effects of these mutations yielded strikingly similar I(Ks) "gain-of-function." In Chinese Hamster Ovary (CHO) cells, coexpression of mutant KCNQ1 with its ancillary subunit KCNE1 generated approximately 3-fold larger currents that activated much faster than wild-type (WT)-I(Ks). Application of the WT NPPA peptide fragment produced similar changes in WT-I(Ks), and these were exaggerated with the mutant NPPA S64R peptide fragment. Anantin, a competitive ANP receptor antagonist, completely inhibited the changes in I(Ks) gating observed with NPPA S64R. Computational simulations identified accelerated transitions into open states as the mechanism for variant I(Ks) gating. Incorporating these I(Ks) changes into computed human atrial action potentials (AP) resulted in 37% shortening (120 vs. 192 ms at 300 ms cycle length), reflecting loss of the phase II dome which is dependent on L-type calcium channel current. We found striking functional similarities due to mutations in KCNQ1 and NPPA genes which led to I(Ks) "gain-of-function", atrial AP shortening, and consequently altered calcium current as a common mechanism between diverse familial AF syndromes.
Copyright 2009 Elsevier Inc. All rights reserved.
Figures
References
-
- Go AS, Hylek EM, Phillips KA, Chang Y, Henault LE, Selby JV, et al. Prevalence of diagnosed atrial fibrillation in adults: national implications for rhythm management and stroke prevention: the AnTicoagulation and Risk Factors in Atrial Fibrillation (ATRIA) Study. JAMA. 2001;285(18):2370–5. - PubMed
-
- Lloyd-Jones DM, Wang TJ, Leip EP, Larson MG, Levy D, Vasan RS, et al. Lifetime risk for development of atrial fibrillation: the Framingham Heart Study. Circulation. 2004;110(9):1042–6. - PubMed
-
- Wolf PA, Abbott RD, Kannel WB. Atrial fibrillation as an independent risk factor for stroke: the Framingham Study. Stroke. 1991;22(8):983–8. - PubMed
-
- Benjamin EJ, Wolf PA, D'Agostino RB, Silbershatz H, Kannel WB, Levy D. Impact of atrial fibrillation on the risk of death: the Framingham Heart Study. Circulation. 1998;98(10):946–52. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
