

Two-site phosphorylation of EPRS coordinates multimodal regulation of noncanonical translational control activity
- PMID: 19647514
- PMCID: PMC2752289
- DOI: 10.1016/j.molcel.2009.05.028
Two-site phosphorylation of EPRS coordinates multimodal regulation of noncanonical translational control activity
Abstract
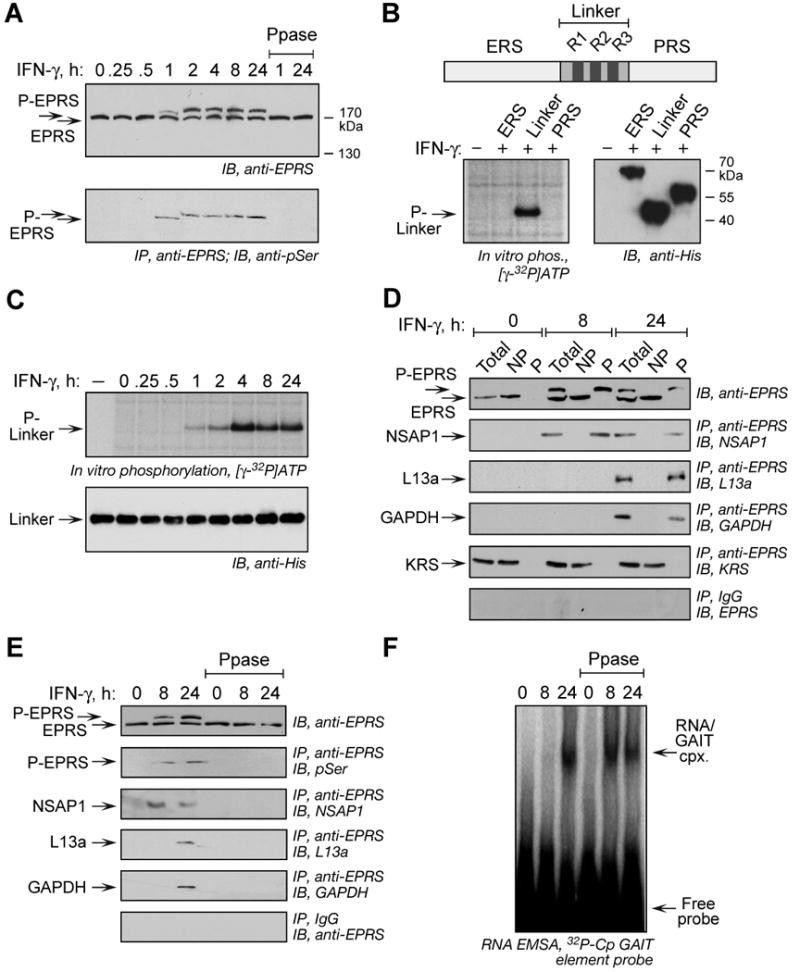
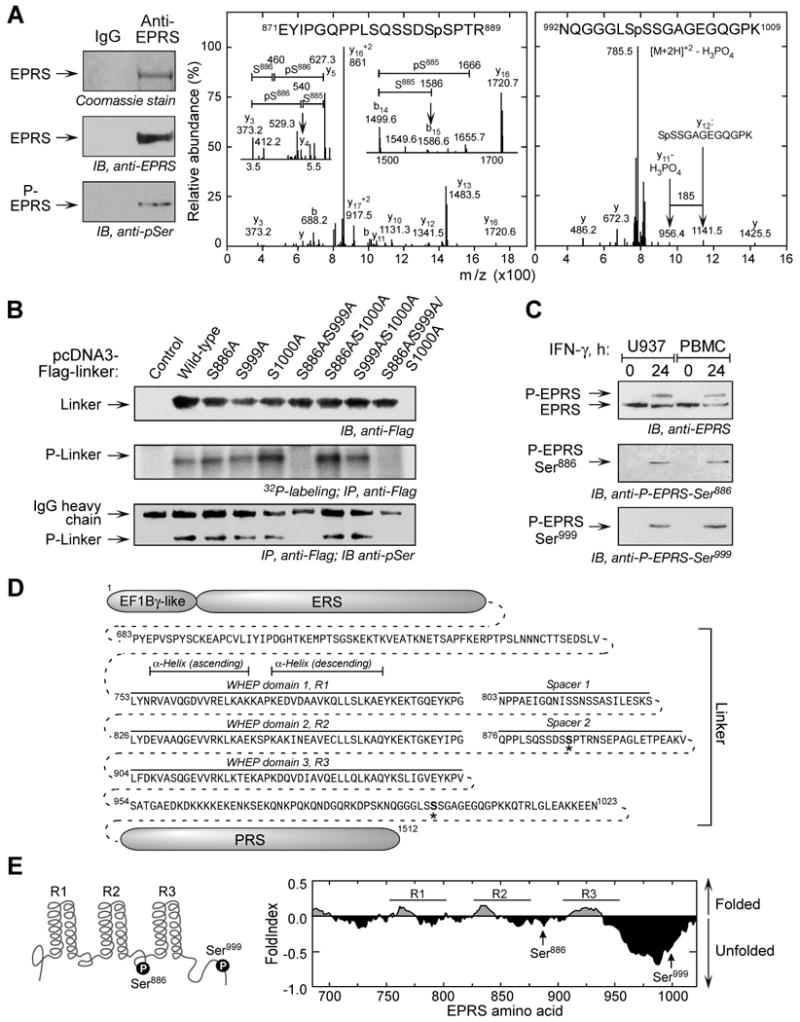
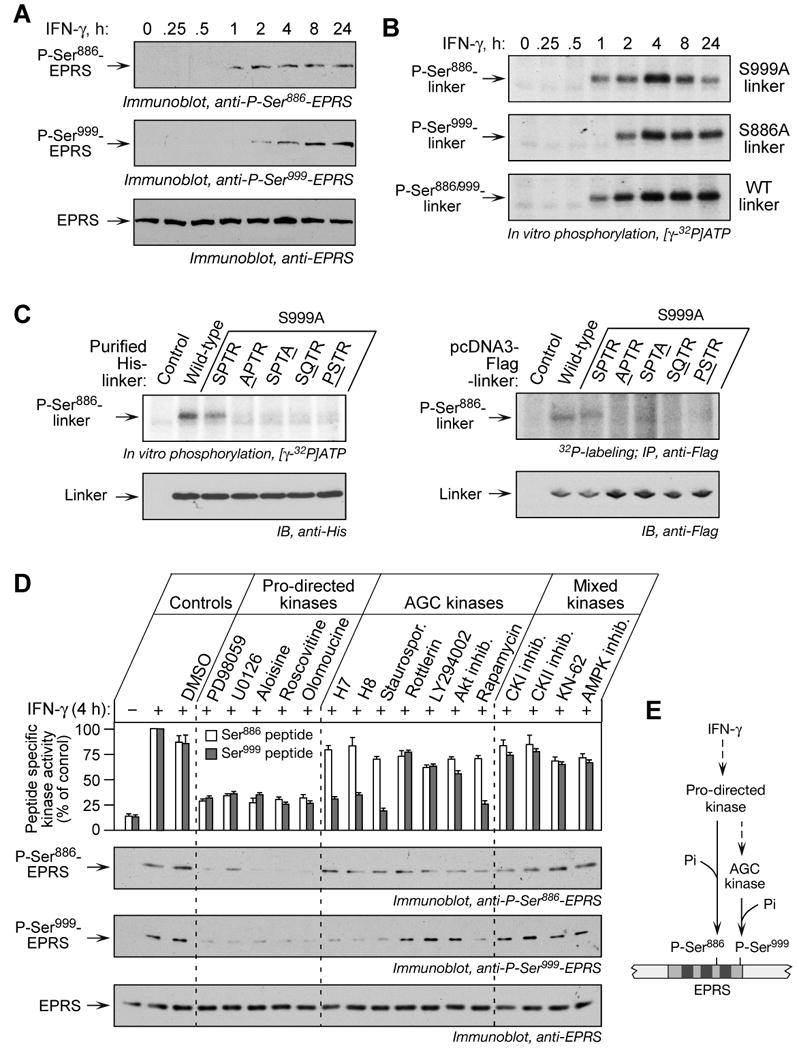
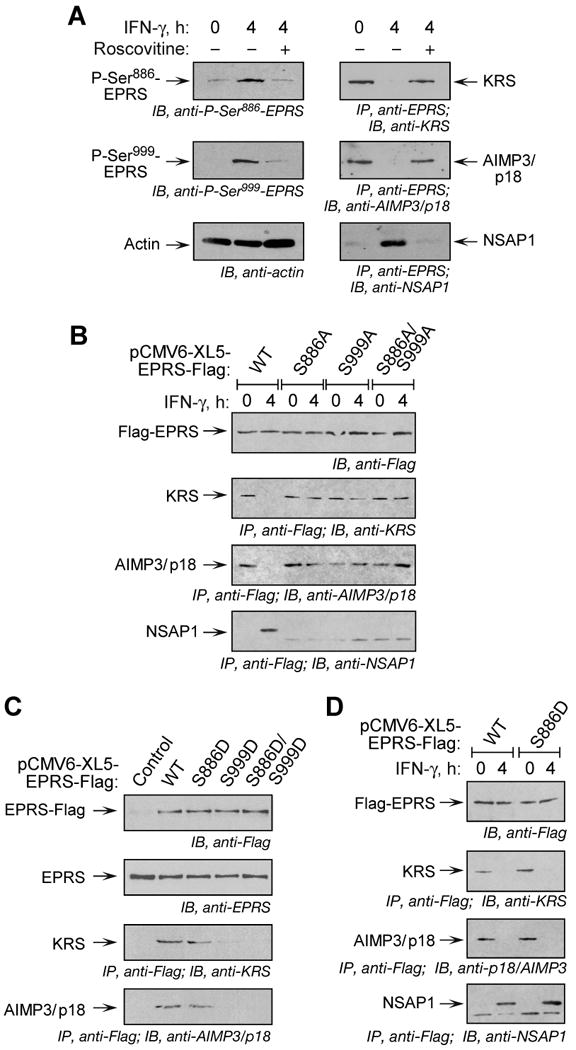
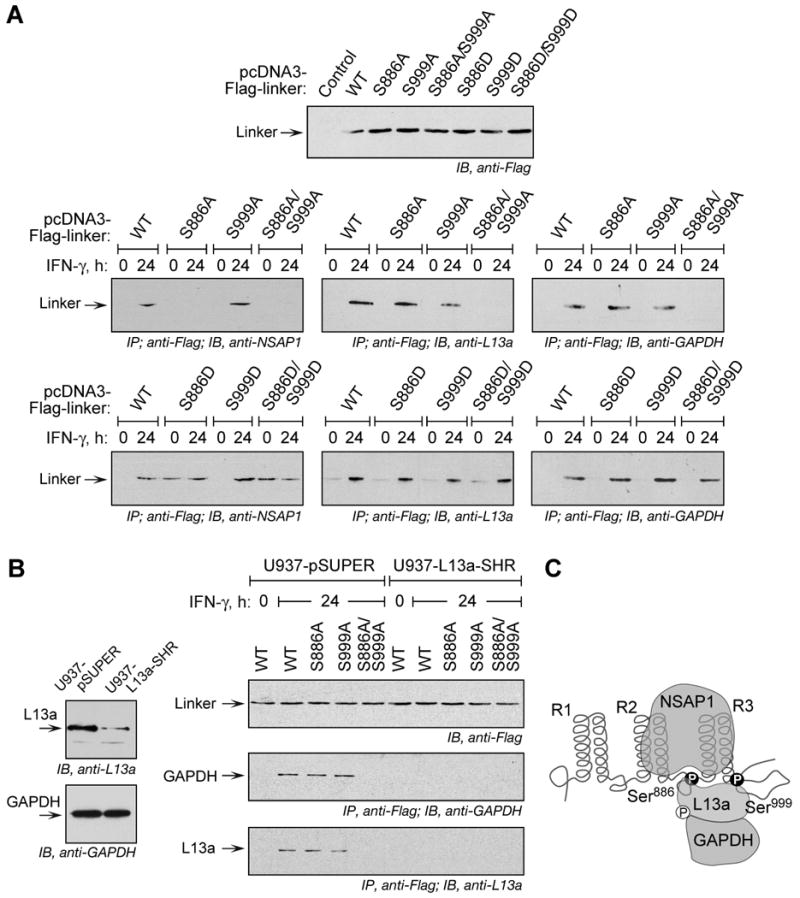
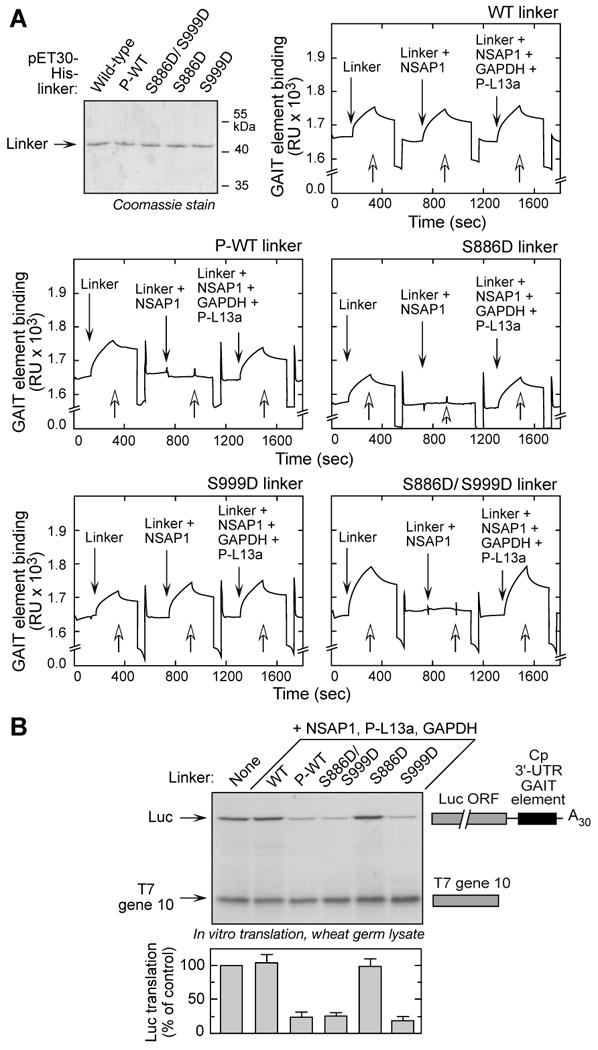
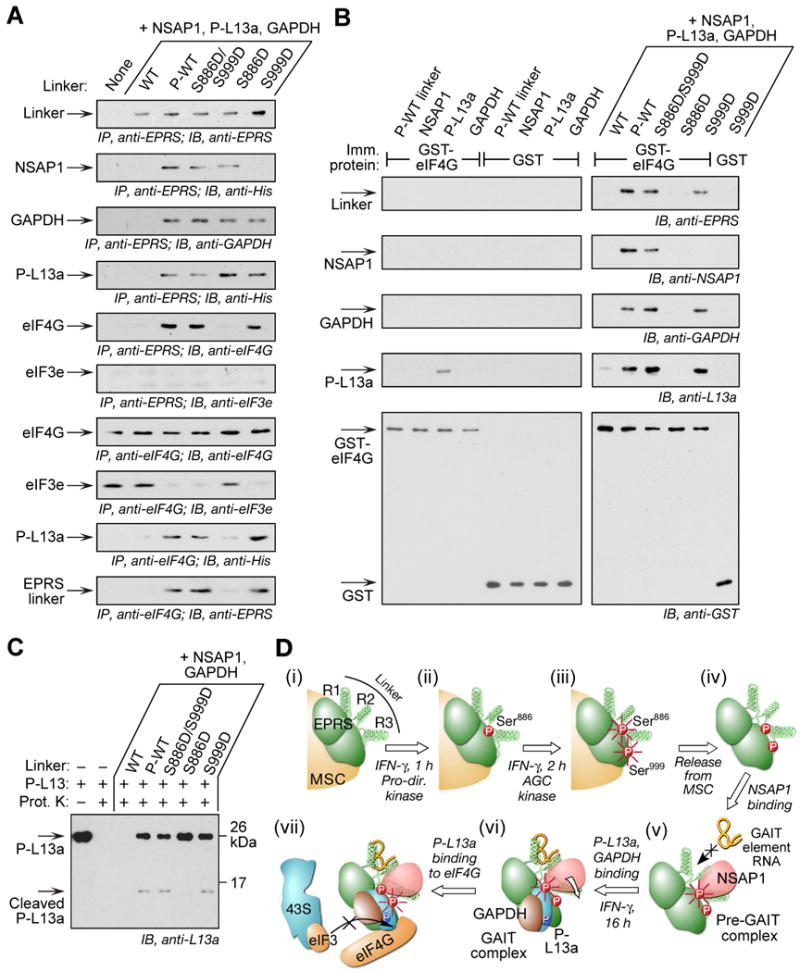
Glutamyl-prolyl tRNA synthetase (EPRS) is a component of the heterotetrameric gamma-interferon-activated inhibitor of translation (GAIT) complex that binds 3'UTR GAIT elements in multiple interferon-gamma (IFN-gamma)-inducible mRNAs and suppresses their translation. Here, we elucidate the specific EPRS phosphorylation events that regulate GAIT-mediated gene silencing. IFN-gamma induces sequential phosphorylation of Ser(886) and Ser(999) in the noncatalytic linker connecting the synthetase cores. Phosphorylation of both sites is essential for EPRS release from the parent tRNA multisynthetase complex. Ser(886) phosphorylation is required for the interaction of NSAP1, which blocks EPRS binding to target mRNAs. The same phosphorylation event induces subsequent binding of ribosomal protein L13a and GAPDH and restores mRNA binding. Finally, Ser(999) phosphorylation directs the formation of a functional GAIT complex that binds initiation factor eIF4G and represses translation. Thus, two-site phosphorylation provides structural and functional pliability to EPRS and choreographs the repertoire of activities that regulates inflammatory gene expression.
Figures
References
-
- Amanchy R, Periaswamy B, Mathivanan S, Reddy R, Tattikota SG, Pandey A. A curated compendium of phosphorylation motifs. Nat Biotechnol. 2007;25:285–286. - PubMed
-
- Berg BH. The early influence of 17-β-oestradiol on 17 aminoacyl-tRNA synthetases of mouse uterus and liver. Phosphorylation as a regulation mechanism. Biochim Biophys Acta. 1977;479:152–171. - PubMed
-
- Casnellie JE. Assay of protein kinases using peptides with basic residues for phosphocellulose binding. Methods Enzymol. 1991;200:115–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
