

Inflammasome-dependent caspase-1 activation in cervical epithelial cells stimulates growth of the intracellular pathogen Chlamydia trachomatis
- PMID: 19648107
- PMCID: PMC2785367
- DOI: 10.1074/jbc.M109.026823
Inflammasome-dependent caspase-1 activation in cervical epithelial cells stimulates growth of the intracellular pathogen Chlamydia trachomatis
Abstract
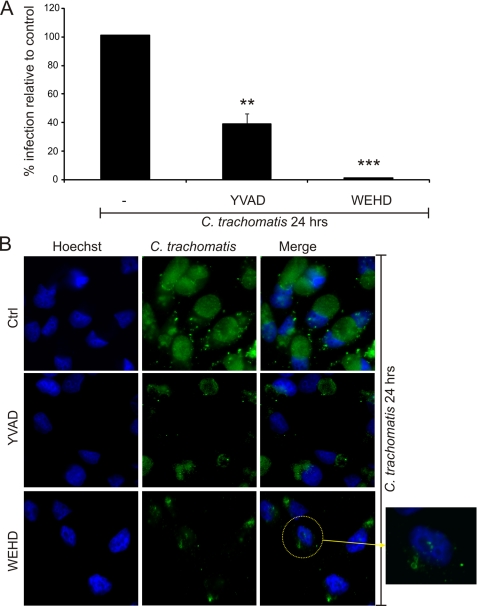
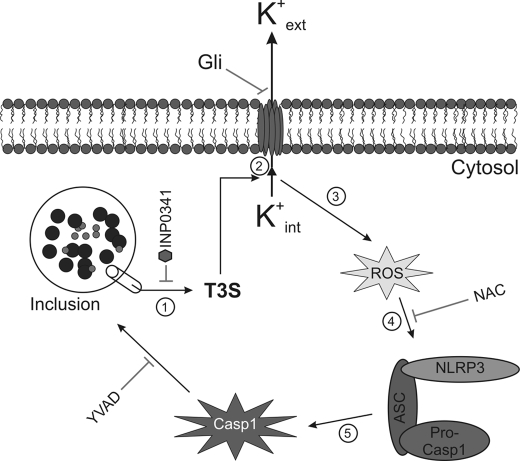
Inflammasomes have been extensively characterized in monocytes and macrophages, but not in epithelial cells, which are the preferred host cells for many pathogens. Here we show that cervical epithelial cells express a functional inflammasome. Infection of the cells by Chlamydia trachomatis leads to activation of caspase-1, through a process requiring the NOD-like receptor family member NLRP3 and the inflammasome adaptor protein ASC. Secretion of newly synthesized virulence proteins from the chlamydial vacuole through a type III secretion apparatus results in efflux of K(+) through glibenclamide-sensitive K(+) channels, which in turn stimulates production of reactive oxygen species. Elevated levels of reactive oxygen species are responsible for NLRP3-dependent caspase-1 activation in the infected cells. In monocytes and macrophages, caspase-1 is involved in processing and secretion of pro-inflammatory cytokines such as interleukin-1beta. However, in epithelial cells, which are not known to secrete large quantities of interleukin-1beta, caspase-1 has been shown previously to enhance lipid metabolism. Here we show that, in cervical epithelial cells, caspase-1 activation is required for optimal growth of the intracellular chlamydiae.
Figures
References
-
- Gerbase A. C., Rowley J. T., Mertens T. E. (1998) Lancet 351, 2–4 - PubMed
-
- Miller W. C., Ford C. A., Morris M., Handcock M. S., Schmitz J. L., Hobbs M. M., Cohen M. S., Harris K. M., Udry J. R. (2004) JAMA 291, 2229–2236 - PubMed
-
- Schachter J. (1999) in Chlamydia: Intracellular Biology, Pathogenesis, and Immunity ( Stephens R. S. ed) pp. 139–170, American Society for Microbiology, Washington, DC
-
- Belland R., Ojcius D. M., Byrne G. I. (2004) Nat. Rev. Microbiol. 2, 530–531 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
