

Vitamin D and differentiation in cancer
- PMID: 19650715
- PMCID: PMC2820234
- DOI: 10.1080/10408360902982128
Vitamin D and differentiation in cancer
Abstract
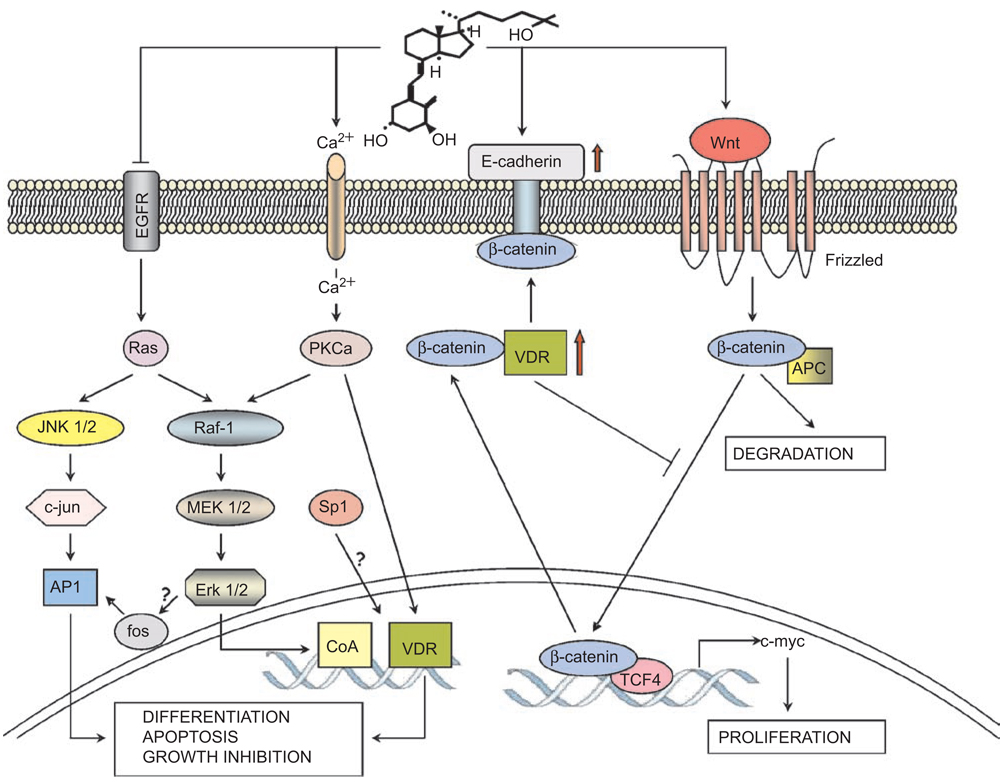
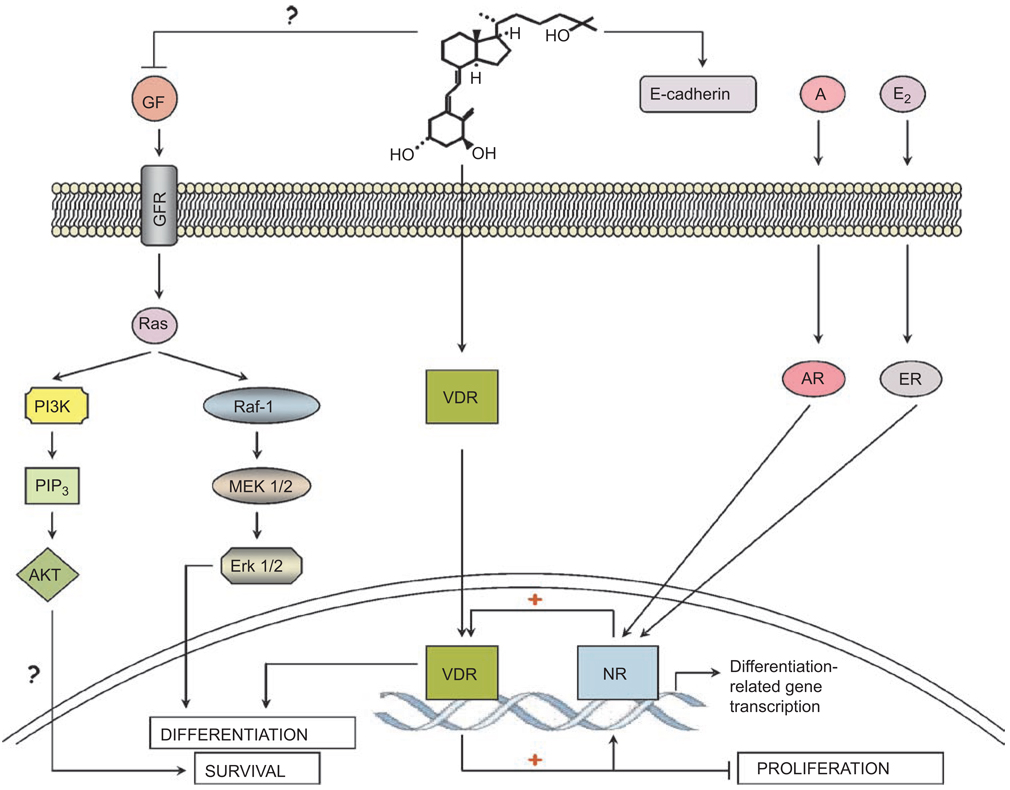
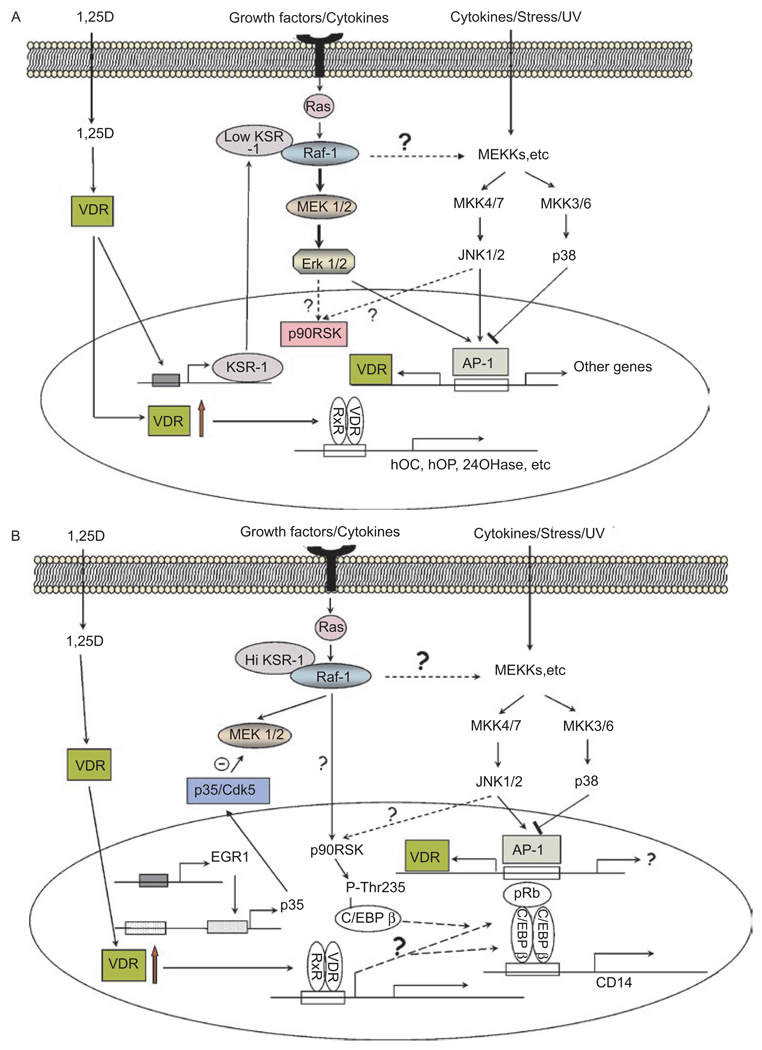
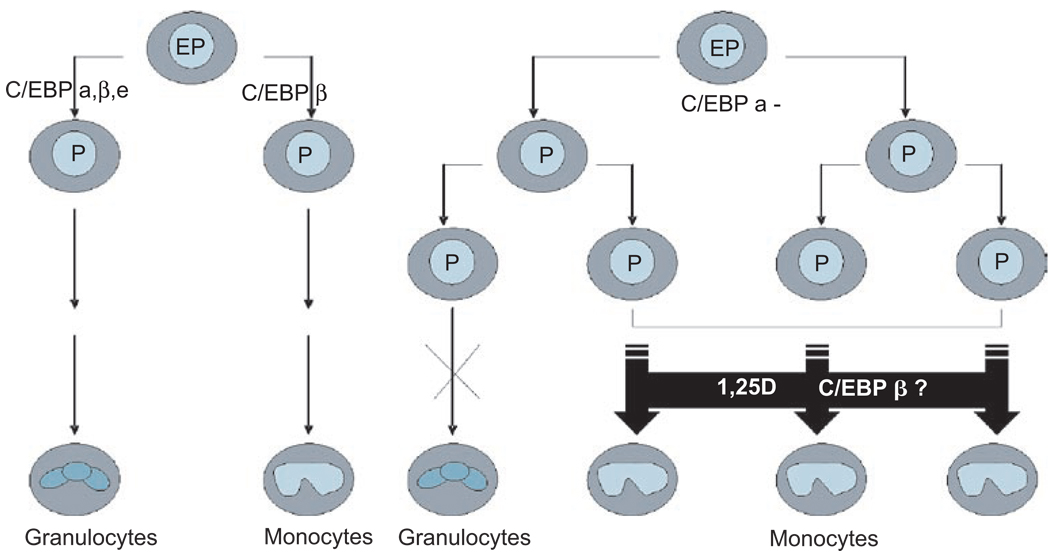
This paper reviews the current understanding of the vitamin D-induced differentiation of neoplastic cells, which results in the generation of cells that acquire near-normal, mature phenotype. Examples of the criteria by which differentiation is recognized in each cell type are provided, and only those effects of 1alpha,25-dihydroxyvitamin D(3) (1,25D) on cell proliferation and survival that are associated with the differentiation process are emphasized. The existing knowledge, often fragmentary, of the signaling pathways that lead to vitamin D-induced differentiation of colon, breast, prostate, squamous cell carcinoma, osteosarcoma, and myeloid leukemia cancer cells is outlined. The important distinctions between the different mechanisms of 1,25D-induced differentiation that are cell-type and cell-context specific are pointed out where known. There is a considerable body of evidence that the principal human cancer cells can be suitable candidates for chemoprevention or differentiation therapy with vitamin D. However, further studies are needed to fully understand the underlying mechanisms in order to improve the therapeutic approaches.
Figures
References
-
- Smithers DW. Maturation in human tumours. Lancet. 1969;2:949–952. - PubMed
-
- Chen SJ, Zhu YJ, Tong JH, Dong S, Huang W, Chen Y, Xiang WM, Zhang L, Li XS, Qian GQ. Rearrangements in the second intron of the RARalfa gene are present in a large majority of patients with acute promyelocytic leukemia and are used as molecular marker for retinoic acid-induced leukemic cell differentiation. Blood. 1991;78:2696–2701. - PubMed
-
- Degos L, Wang ZY. All trans retinoic acid in acute promyelocytic leukemia. Oncogene. 2001;20:7140–7145. - PubMed
-
- Schlenk RF, Frohling S, Hartmann F, Fischer JT, Glasmacher A, del Valle F, Grimminger W, Gotze K, Waterhouse C, Schoch R, Pralle H, Mergenthaler HG, Hensel M, Koller E, Kirchen H, Preiss J, Salwender H, Biedermann HG, Kremers S, Griesinger F, Benner A, Addamo B, Dohner K, Haas R, Dohner H. Phase III study of all-trans retinoic acid in previously untreated patients 61 years or older with acute myeloid leukemia. Leukemia. 2004;18:1798–1803. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources