

Review
doi: 10.1016/j.ymeth.2009.07.005.
Epub 2009 Aug 3.
Identification of catalytic metal ion ligands in ribozymes
Affiliations
- PMID: 19651216
- PMCID: PMC3470912
- DOI: 10.1016/j.ymeth.2009.07.005
Item in Clipboard
Review
Identification of catalytic metal ion ligands in ribozymes
Methods.
2009 Oct.
Abstract
Site-bound metal ions participate in the catalytic mechanisms of many ribozymes. Understanding these mechanisms therefore requires knowledge of the specific ligands on both substrate and ribozyme that coordinate these catalytic metal ions. A number of different structural and biochemical strategies have been developed and refined for identifying metal ion binding sites within ribozymes, and for assessing the catalytic contributions of the metal ions bound at those sites. We review these approaches and provide examples of their application, focusing in particular on metal ion rescue experiments and their roles in the construction of the transition state models for the Tetrahymena group I and RNase P ribozymes.
Figures
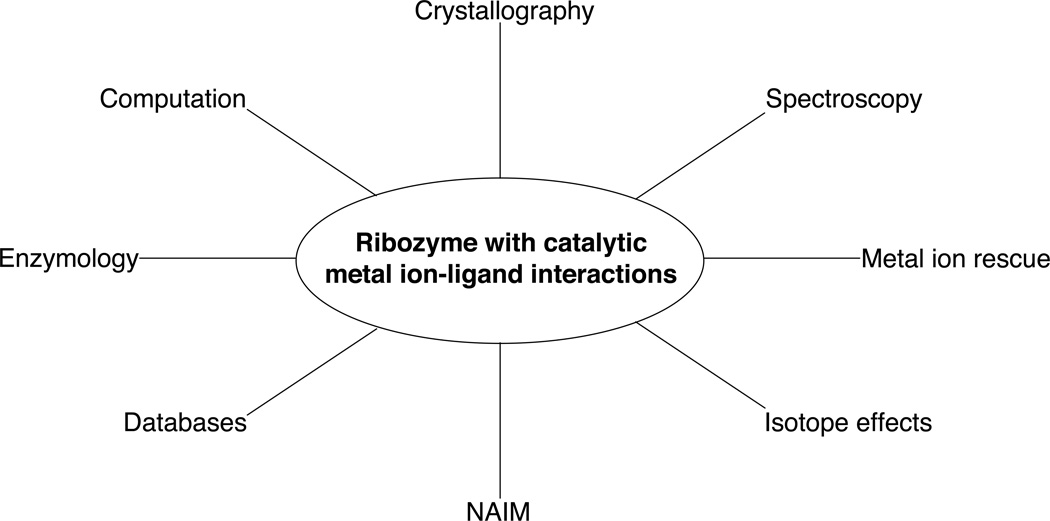
Summary of techniques used to identify catalytic metal ion-ligand interactions within ribozymes.
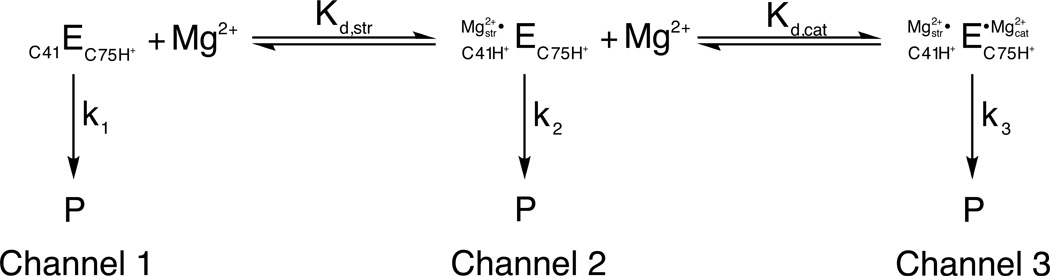
A three-channel scheme describing the Mg2+-dependent and independent pathways for HDV ribozyme catalysis. In 1 M NaCl in the absence of Mg2+, the ribozyme reacts at a basal rate with rate constant k1 (channel 1). Increasing concentrations of Mg2+ accelerate this rate in a log-linear fashion (channel 2) up to a point, after which the rate becomes independent of Mg2+ concentration (channel 3). The Mg2+ ion associated with channel 2 conditions is thought to be primarily structural, binding via inner sphere coordination to a base quadruple involving a protonated C41. Under channel 3 conditions, a second, catalytic, Mg2+ ion binds via outer sphere coordination in the vicinity of the active site. This fully hydrated Mg2+ ion is thought to participate in general acid-base catalysis. [Adapted from (10).]
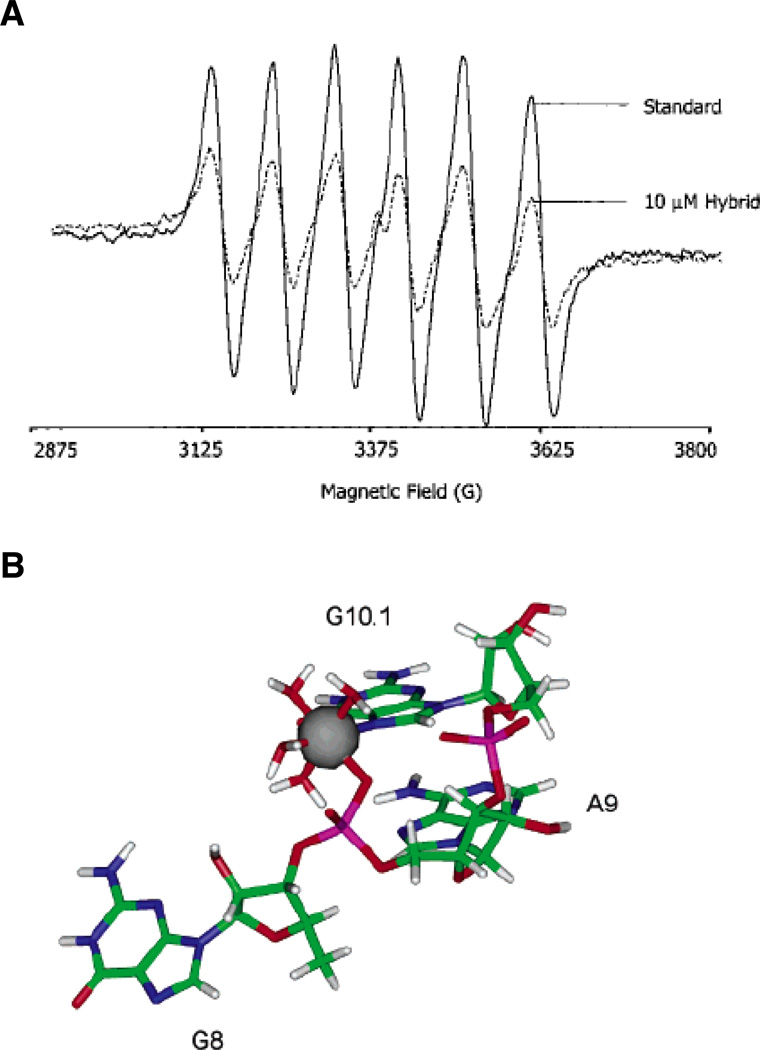
EPR spectroscopy of a Mn2+ binding site within the hammerhead ribozyme. (A) EPR spectra of a 50 µM solution of Mn2+ alone (solid line) and in the presence of 10 µM of an RNA-DNA hybrid hammerhead ribozyme (dotted line). The addition of nucleic acid causes the six-line EPR spectrum of Mn2+ to diminish. (B) Model of the A9/G10.1 Mn2+ binding site in the tertiary-stabilized hammerhead ribozyme, as determined through energy minimization of crystallographic and ESEEM data. The Mn2+ ion coordinates the pro-RP nonbridging oxygen of A9, the N7 of G10.1, and four water molecules. [Taken from (43) and (46), with permission.]
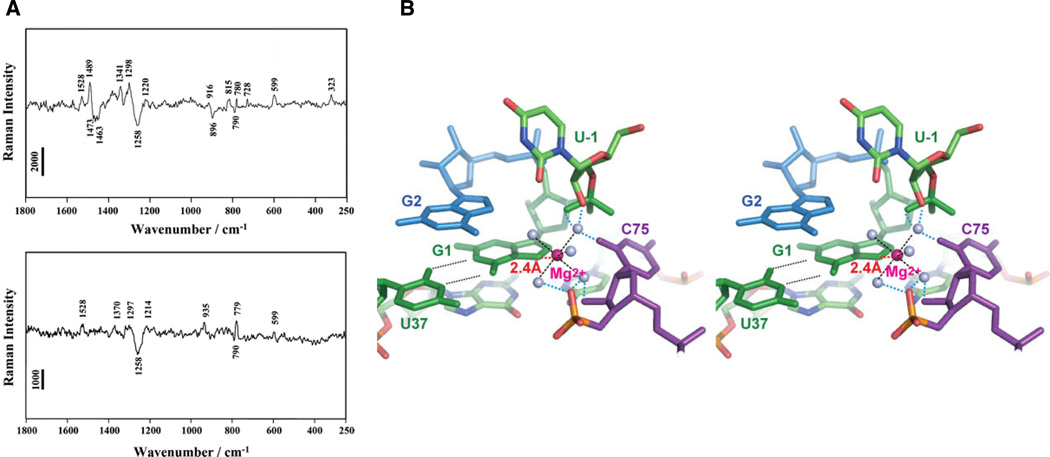
Raman spectroscopic investigation of HDV ribozyme crystals. (A) Raman difference spectra (pH 7.5 – pH 5.0) of crystals of the unmodified HDV ribozyme (top panel) and an HDV ribozyme in which 7-deazaguanosine has replaced the cleavage site G1 (bottom panel). The difference spectrum of the unmodified ribozyme shows peaks at 1489 and 323 cm−1, thought to arise from metal ion coordination of a guanosine N7 and from a single inner sphere Mg2+ hydrate, respectively. Both features are lost when 7-deazaguanosine replaces guanosine at the cleavage site. (B) Stereo model of the HDV ribozyme active site, constructed by combining the Raman spectroscopic results with pre- and post-cleavage crystallographic data. The Mg2+ ion (magenta) directly coordinates the N7 of G1, along with five water molecules (grey spheres). [Taken from (13), with permission.]
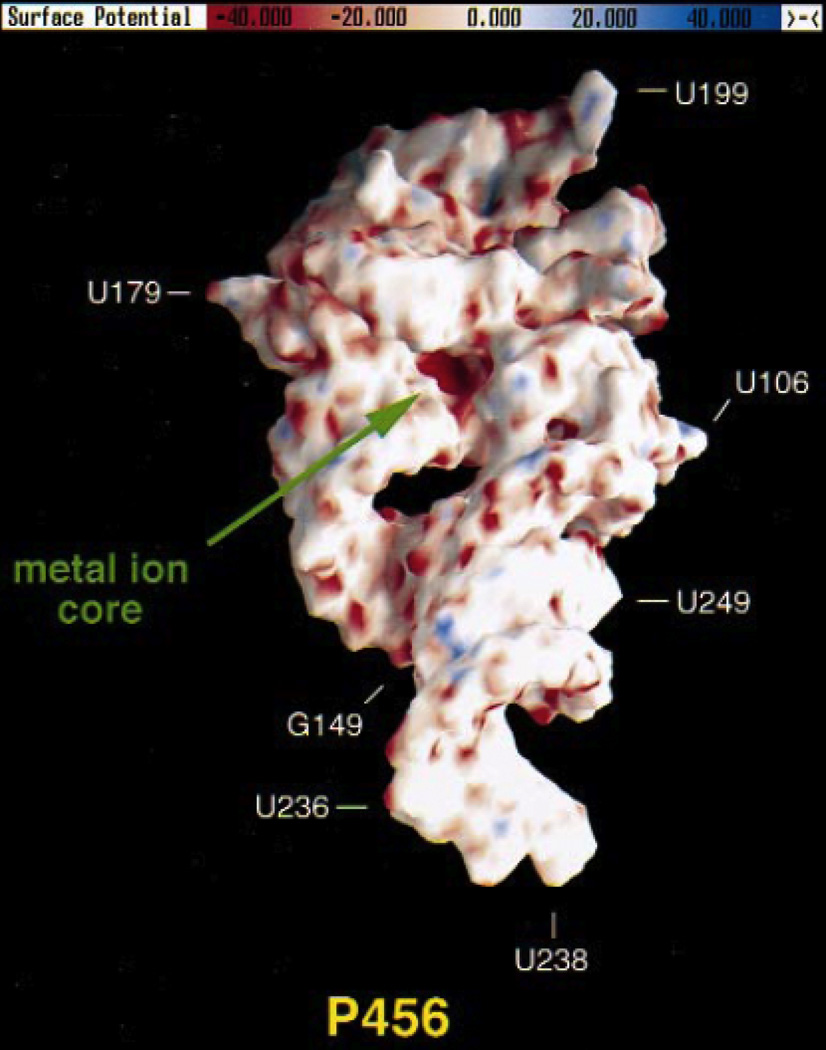
Electrostatic potential surface of the P4–P6 domain from the Tetrahymena group I ribozyme, calculated using the NLPB equation. Regions of negative electrostatic potential appear red, while electropositive regions appear blue. The A-rich bulge that coordinates two Mg2+ ions is located in a pocket of highly negative electrostatic potential (“metal ion core”) with values ranging from −80 to −100 kT/e. [Adapted from (74), with permission.]
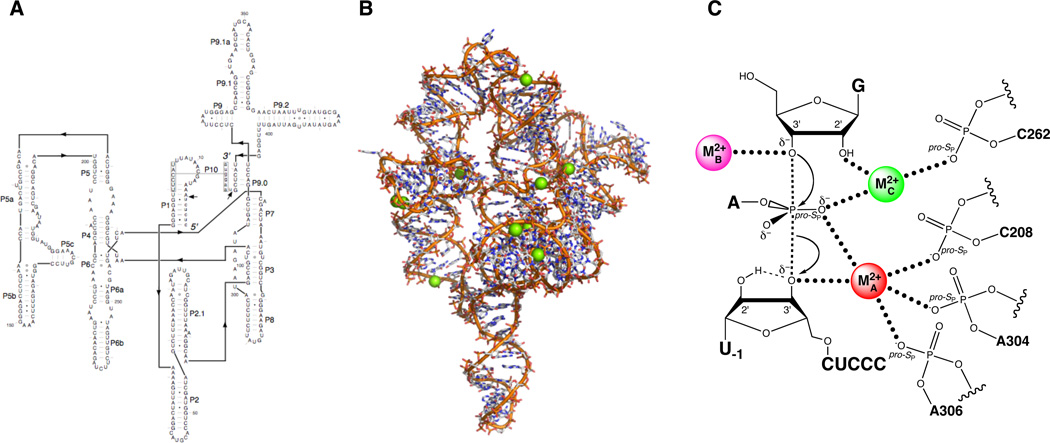
The Tetrahymena group I ribozyme. (A) Secondary structure. (B) Three-dimensional structure at 3.8 Å resolution (PDB file 1X8W). Mg2+ ions are represented as green spheres. (C) Transition state catalytic metal ion-ligand interactions within the Tetrahymena ribozyme active site, as deduced from metal ion rescue experiments.
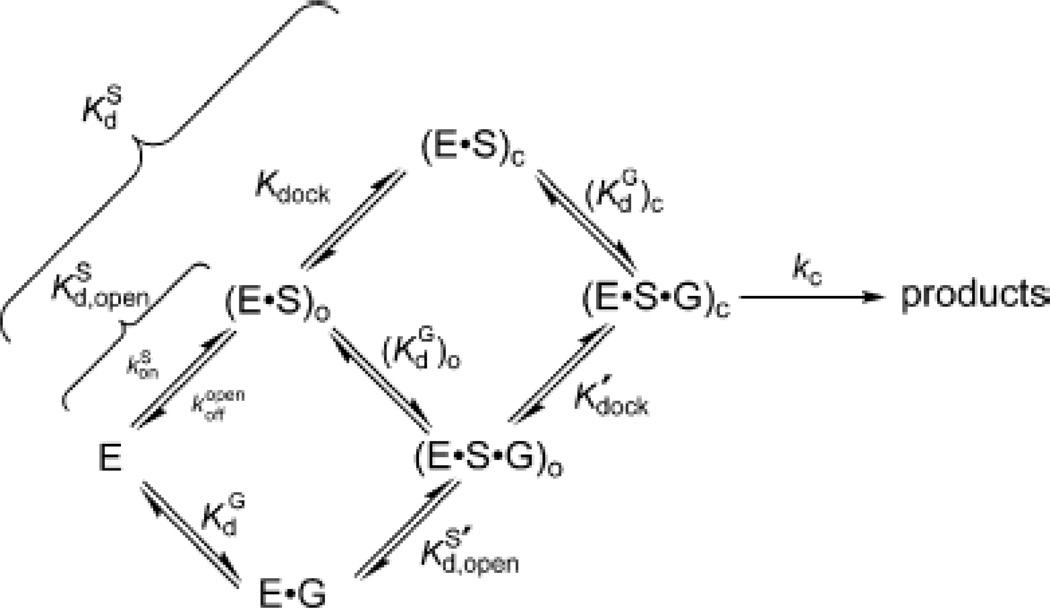
Kinetic model of the Tetrahymena ribozyme reaction. The free enzyme (E) can bind the oligonucleotide substrate (S) to form the open complex, in the presence [(E·S·G)O] or absence [(E·S)O] of guanosine. The closed complex [(E·S·G)C or (E·S)C] forms when the P1 helix carrying the substrate “docks” into the active site of the ribozyme via the formation of tertiary interactions. [Taken from (111).]
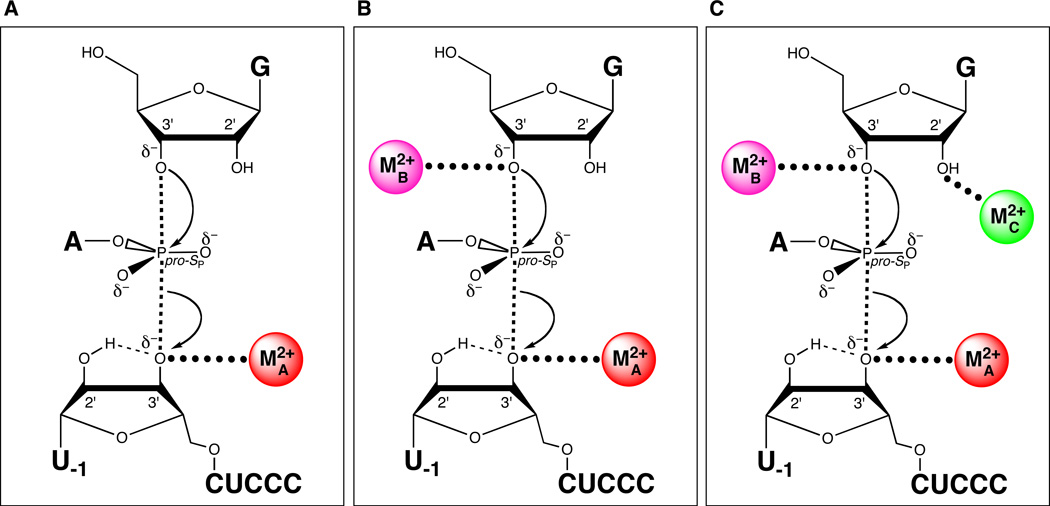
Sequential discovery of three different catalytic metal ion-ligand interactions in the transition state of the Tetrahymena group I ribozyme reaction. (A) MA coordinates the 3’ oxygen of the leaving group. (B) MB coordinates the 3’-OH of the guanosine nucleophile. (C) MC coordinates the 2’-OH of the guanosine nucleophile.
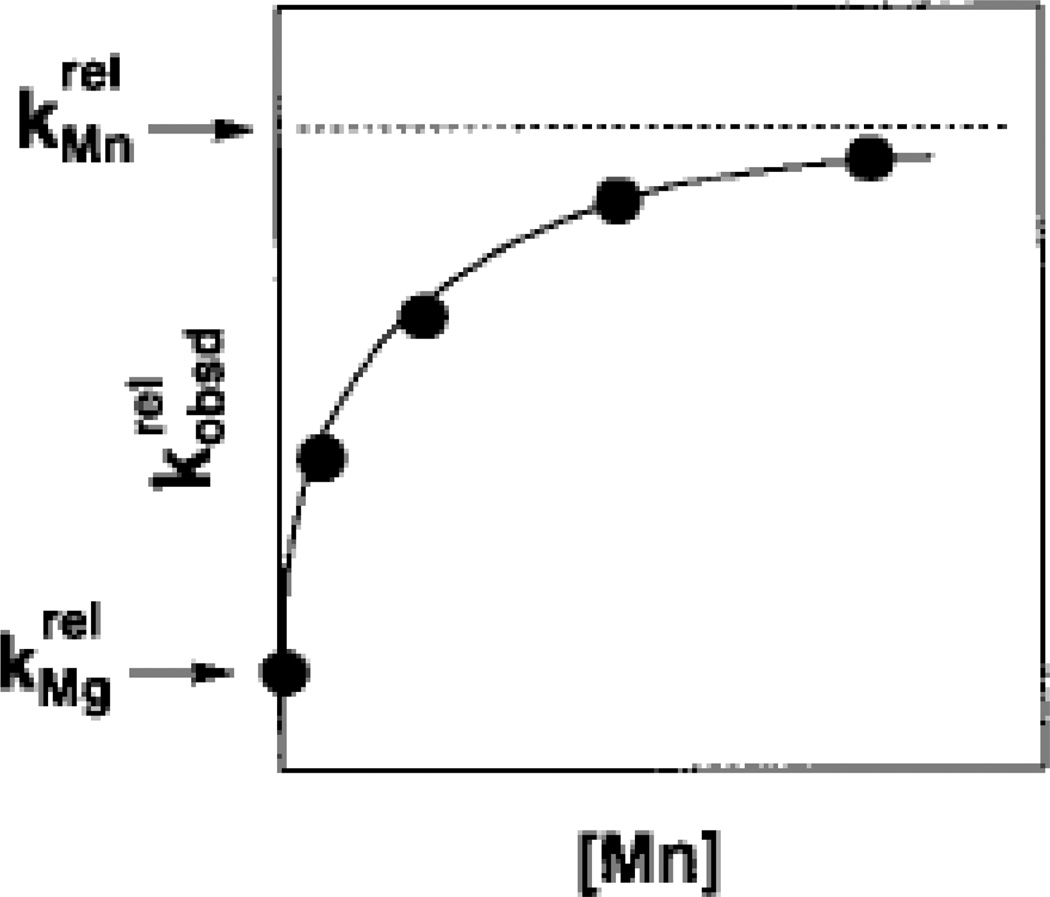
Mn2+ concentration dependence of the reaction of S3’S relative to S3’O () for the reaction shown in Scheme 1. The Mn2+-dependent acceleration of the reaction is associated with a specific affinity (KMn) of the rescuing Mn2+ ion for the free ribozyme. This affinity is a thermodynamic fingerprint for Mn2+ rescue of the 3’ sulfur modification. [Taken from (121).]
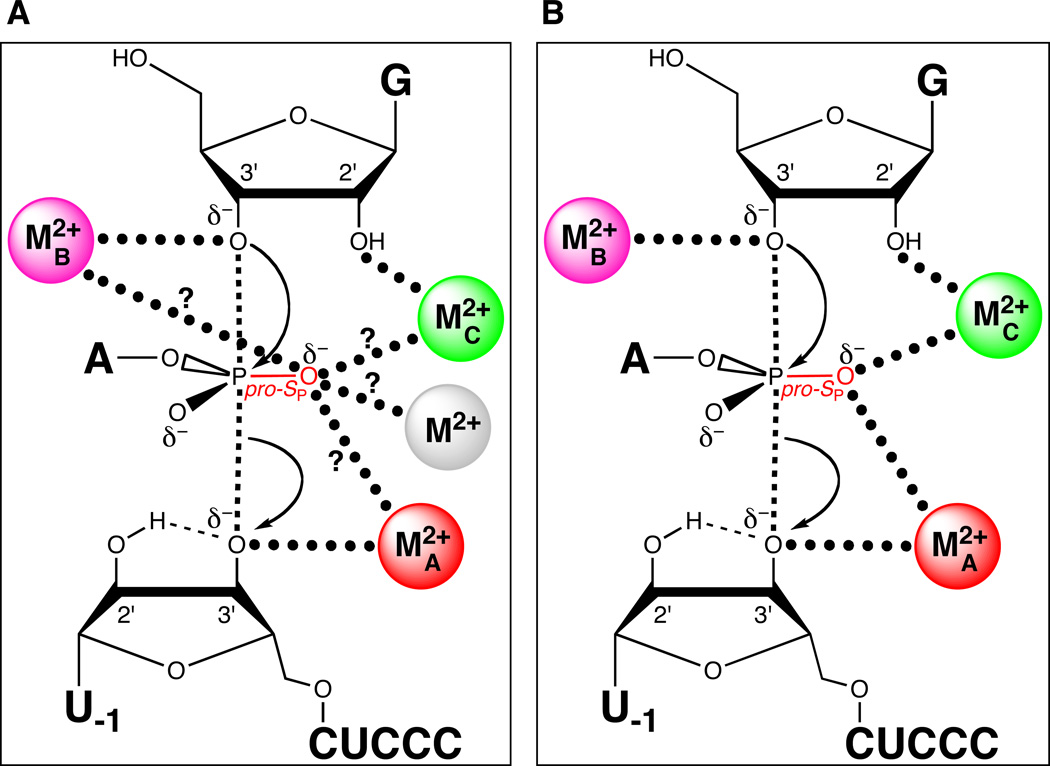
Both MA and MC coordinate the pro-SP nonbridging oxygen of the scissile phosphate. (A) Model showing potential metal ion-ligand interactions involving the pro-SP nonbridging oxygen (question marks). The grey sphere denotes a fourth divalent metal ion, a possibility not excluded from the results in (123). (B) Revised transition state model based on (124). Three metal ions are present in the active site, with metals A and C coordinating the pro-SP nonbridging oxygen of the scissile phosphate.
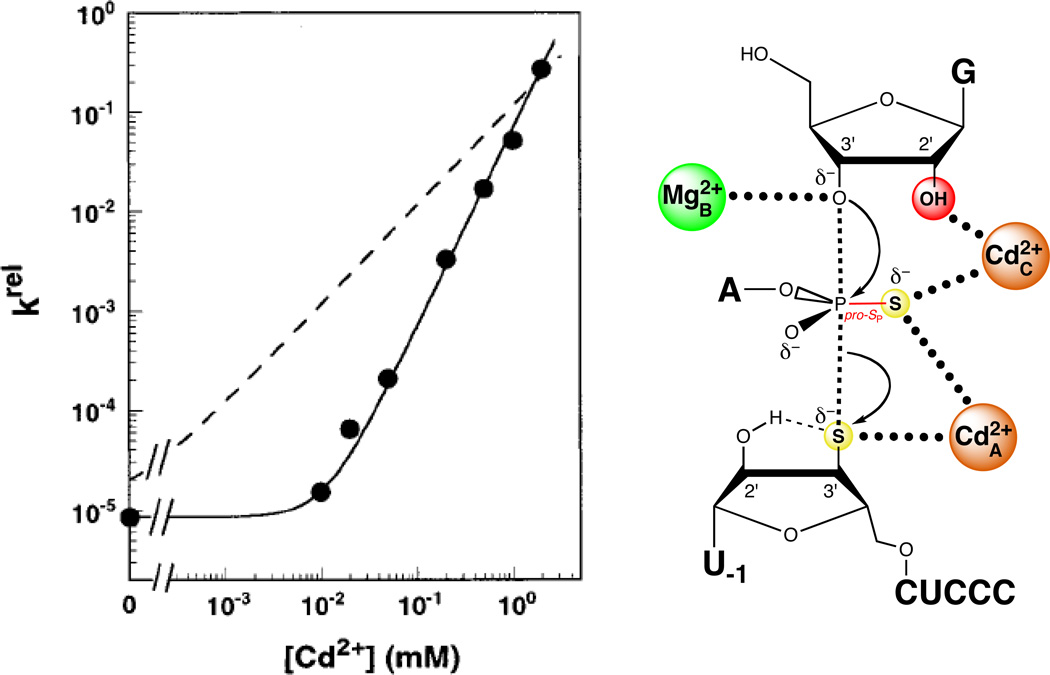
Two Cd2+ ions at sites A and C rescue the reaction of S3’S,P-S by the Tetrahymena ribozyme. The plot at left shows the Cd2+ concentration dependence of krel, the reaction rate constant for S3’S,P-S relative to that of an all-oxygen substrate. The solid line fits the data according to a model in which two Cd2+ ions rescue the reaction, while the dashed line fits the data according to a model in which a single Cd2+ ion mediates rescue. [Data taken from (124).]
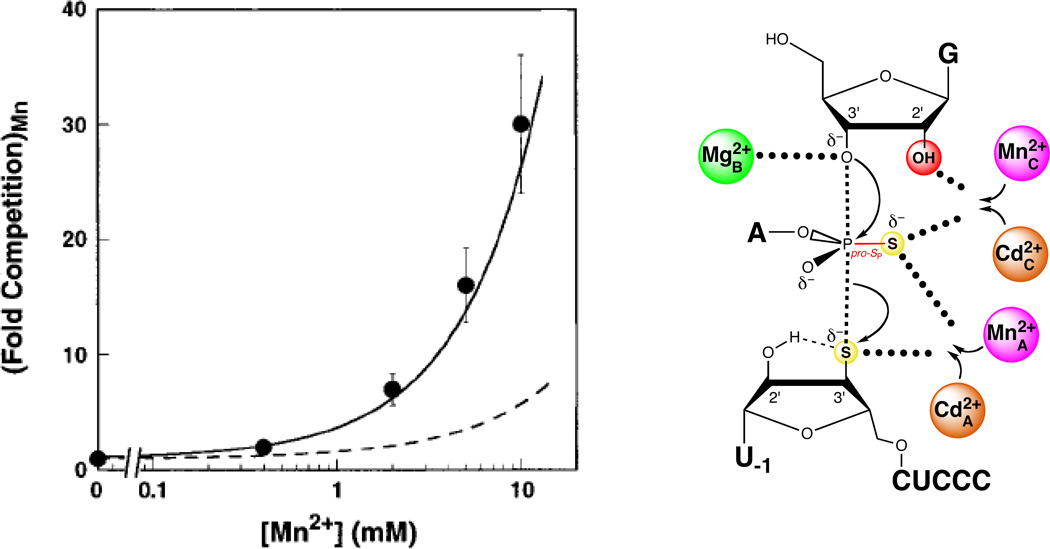
Mn2+ ions compete with Cd2+ ions for sites A and C during rescue of the S3’S,P-S reaction. In the plot at left, the data are represented as filled spheres. The solid line shows the degree of competition expected from Mn2+ ions competing at sites A and C, while the dashed line shows the corresponding model for Mn2+ competition at sites A and B. [Data taken from (124).]
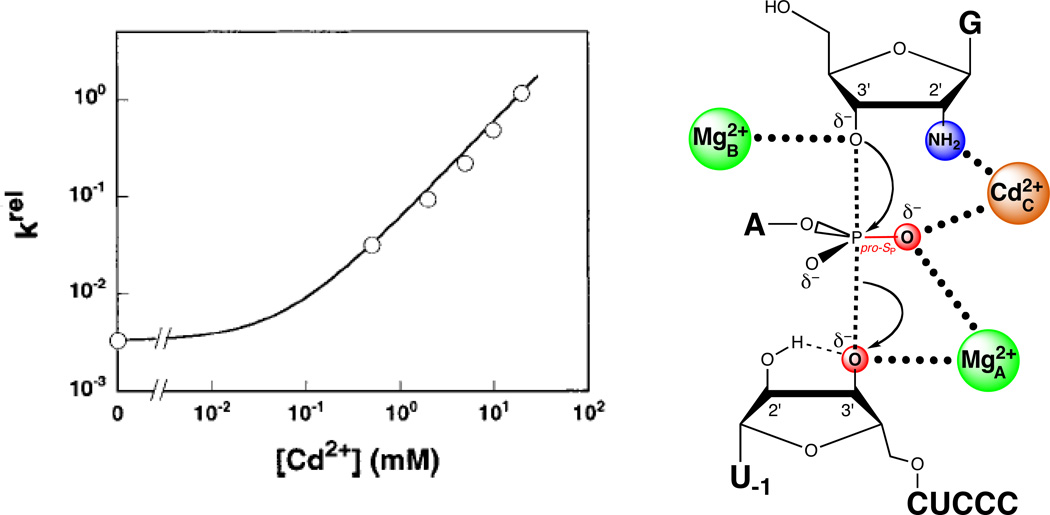
A single Cd2+ ion rescues the reaction in which G2′NH2 replaces guanosine. The plot at left shows the Cd2+ dependence of the rate constant for the reaction involving G2′NH2relative to guanosine for the all-oxygen substrate. The data are fit to a model in which a single Cd2+ ion rescues the reaction. [Data taken from (124).]
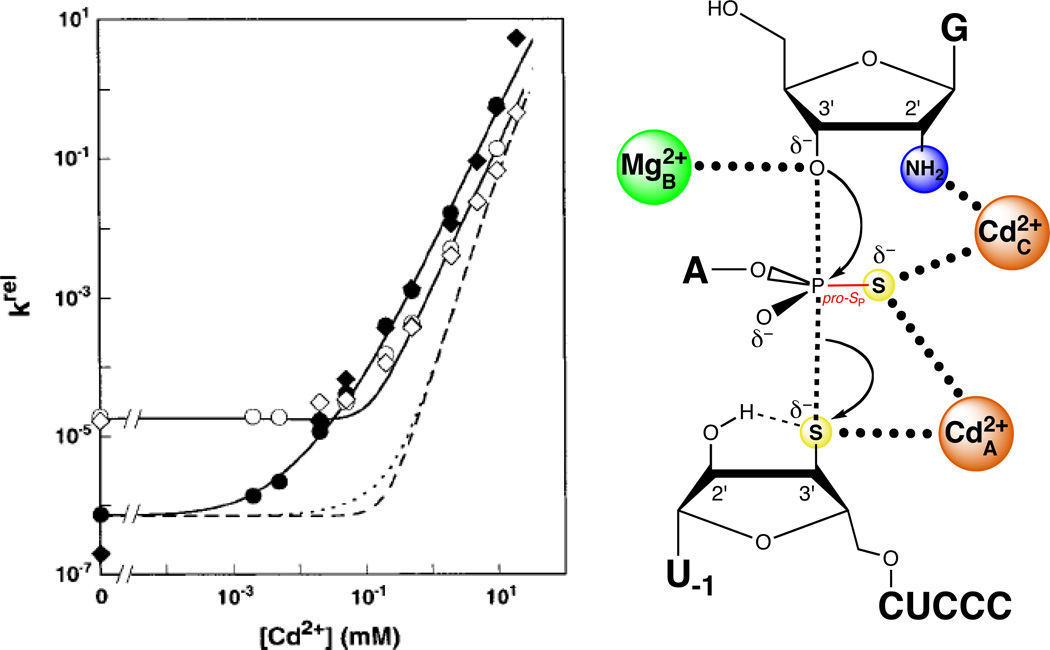
Two Cd2+ ions rescue the reaction of S3’S,P-S with G2′NH2. In the plot at left, the closed symbols denote two independent determinations of the Cd2+ dependence of the reaction of S3’S,P-S with G2′NH2, relative to that of the all-oxygen substrate with guanosine. The open symbols denote two independent determinations of krel for the reaction of S3’S,P-S with guanosine, compared to that of the all-oxygen substrate with guanosine. The solid lines are fits of the data to a model in which two Cd2+ ions rescue the reaction, while the dashed lines are fits in which three Cd2+ ions rescue the reaction. [Data taken from (124).]
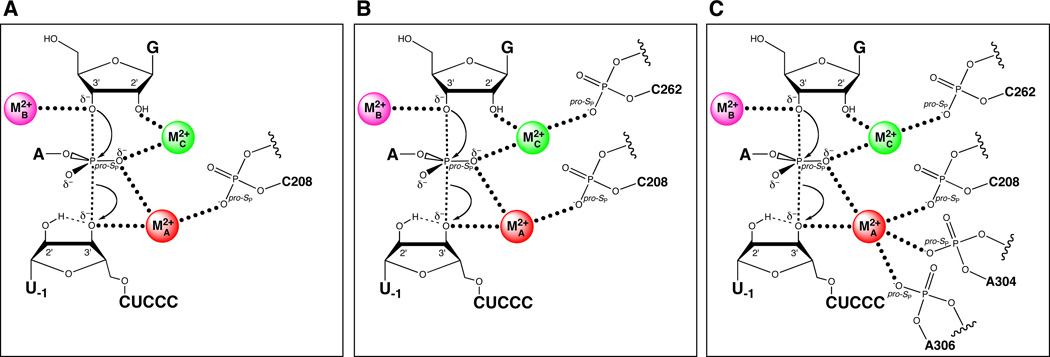
Sequential discovery of ligands within the Tetrahymena ribozyme that position the catalytic metal ions. (A) The pro-SP nonbridging oxygen of C208 coordinates MA. (B) The pro-SP nonbridging oxygen of C262 coordinates MC. (C) Additional ligands for MA include The pro-SP nonbridging oxygens of A304 and A306.
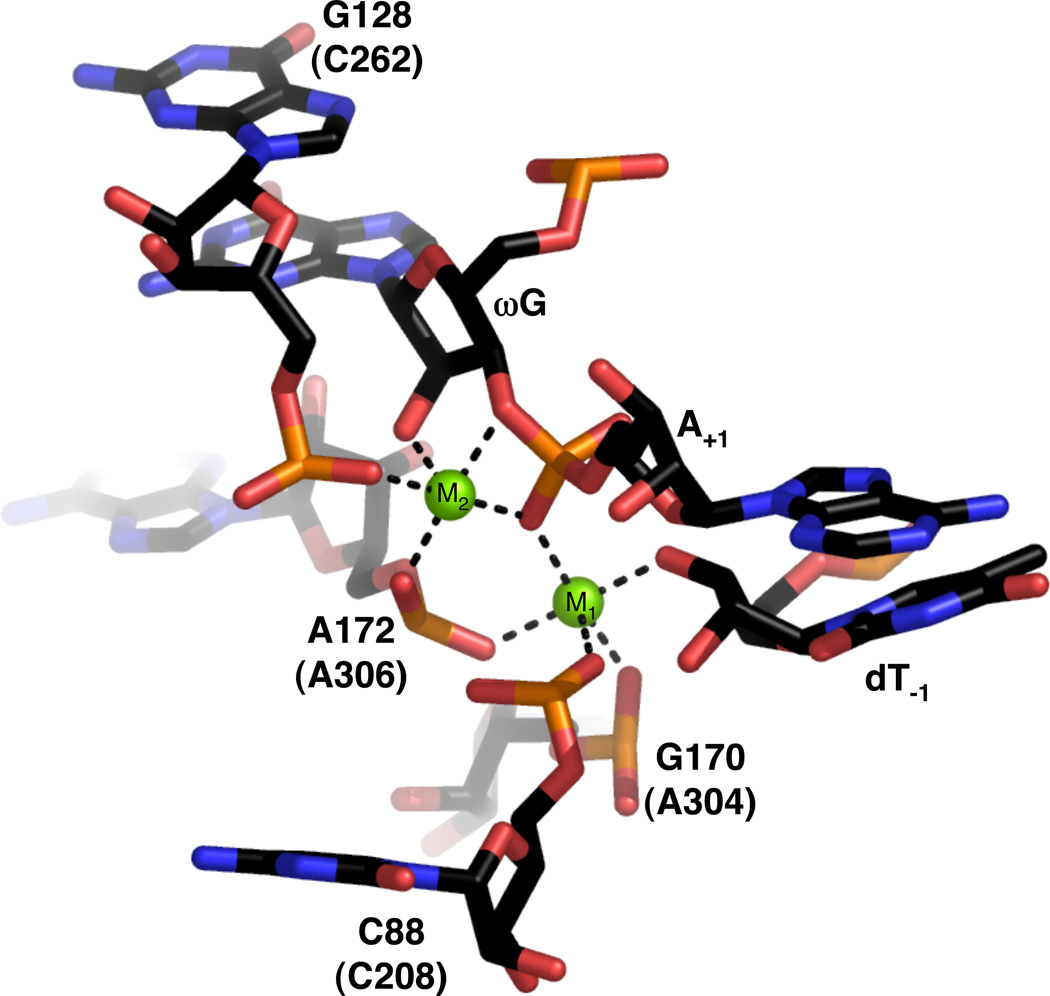
View of the active site of the Azoarcus group I ribozyme (PDB file 1ZZN). Nucleotide numbers in parentheses refer to homologous positions within the Tetrahymena ribozyme. Mg2+ ions are represented as green spheres. Black lines denote putative inner sphere interactions between ribozyme ligands and the metal ions.
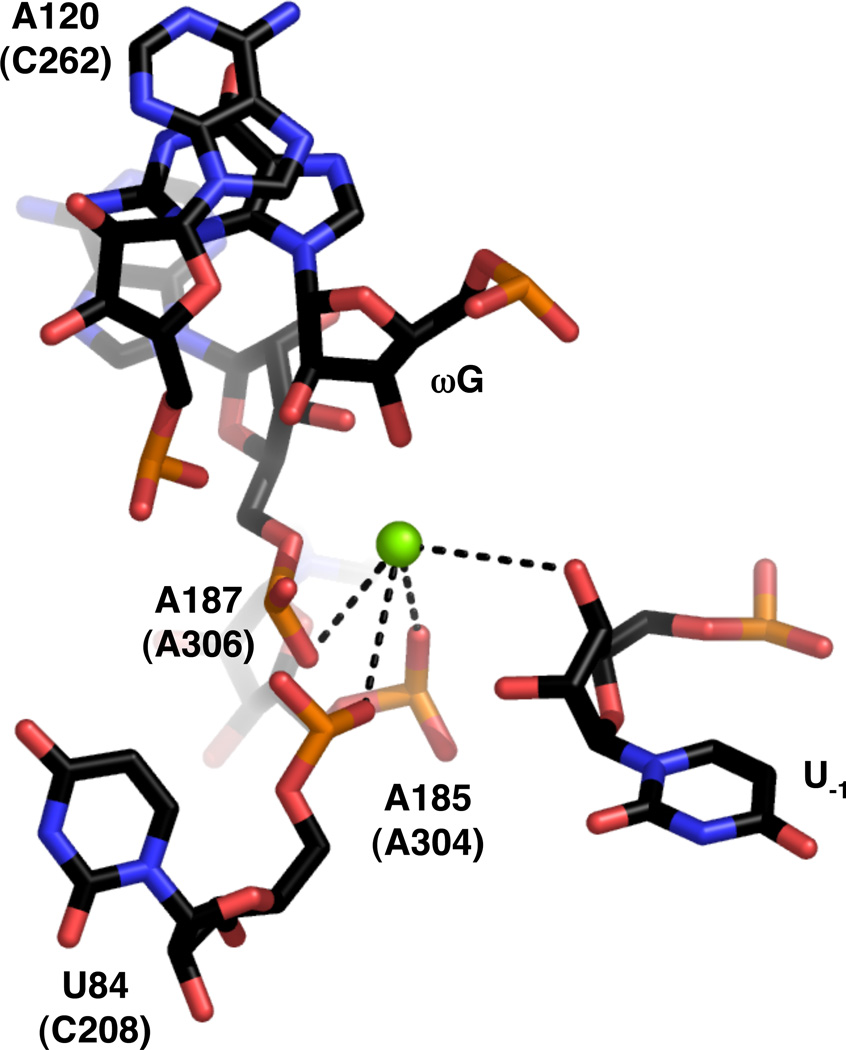
View of the active site of the group I ribozyme derived from bacteriophage Twort (PDB file 1Y0Q). Numbering and colors are as in Figure 16.
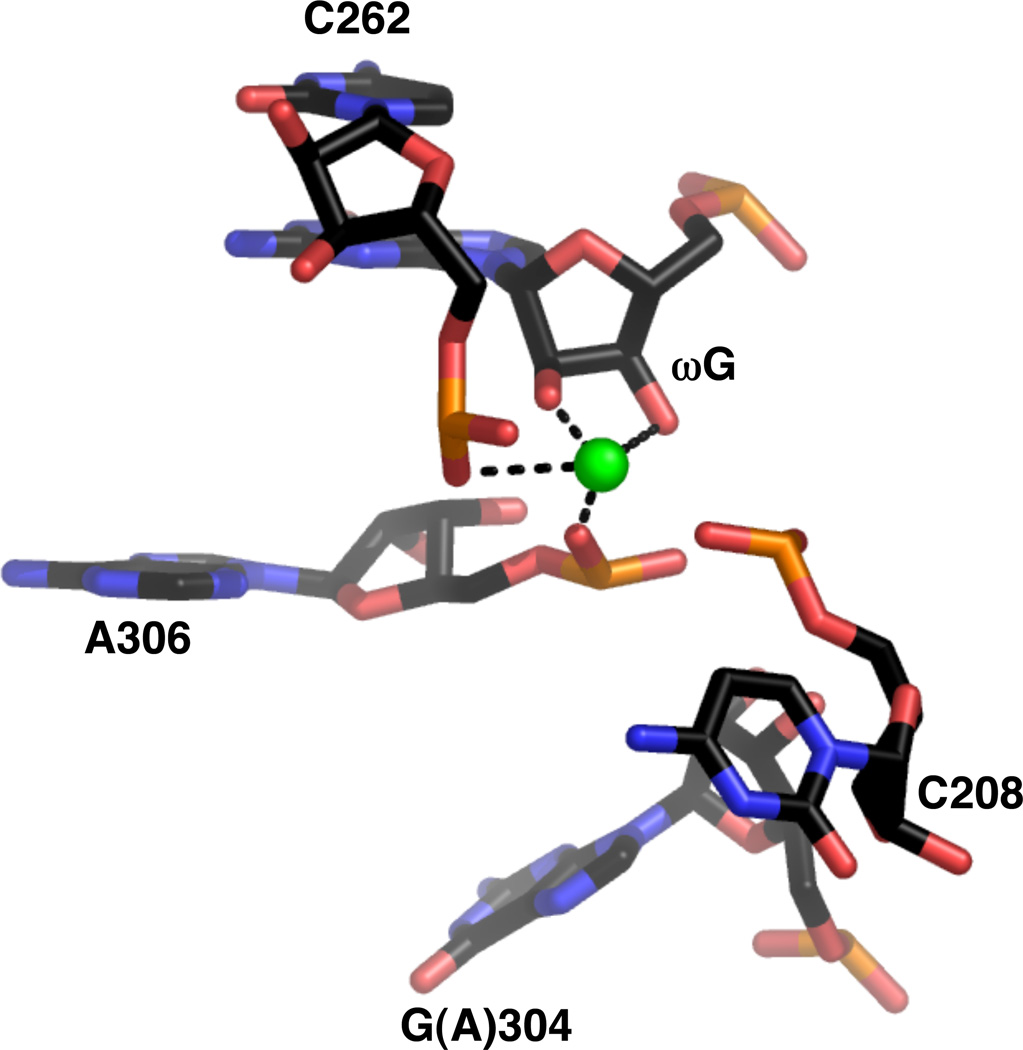
Active site metal ion within the crystal structure of the Tetrahymena ribozyme (PDB file 1X8W). The guanosine at position 304 is one of several mutations introduced based on experiments that selected for thermostable variants of the ribozyme.
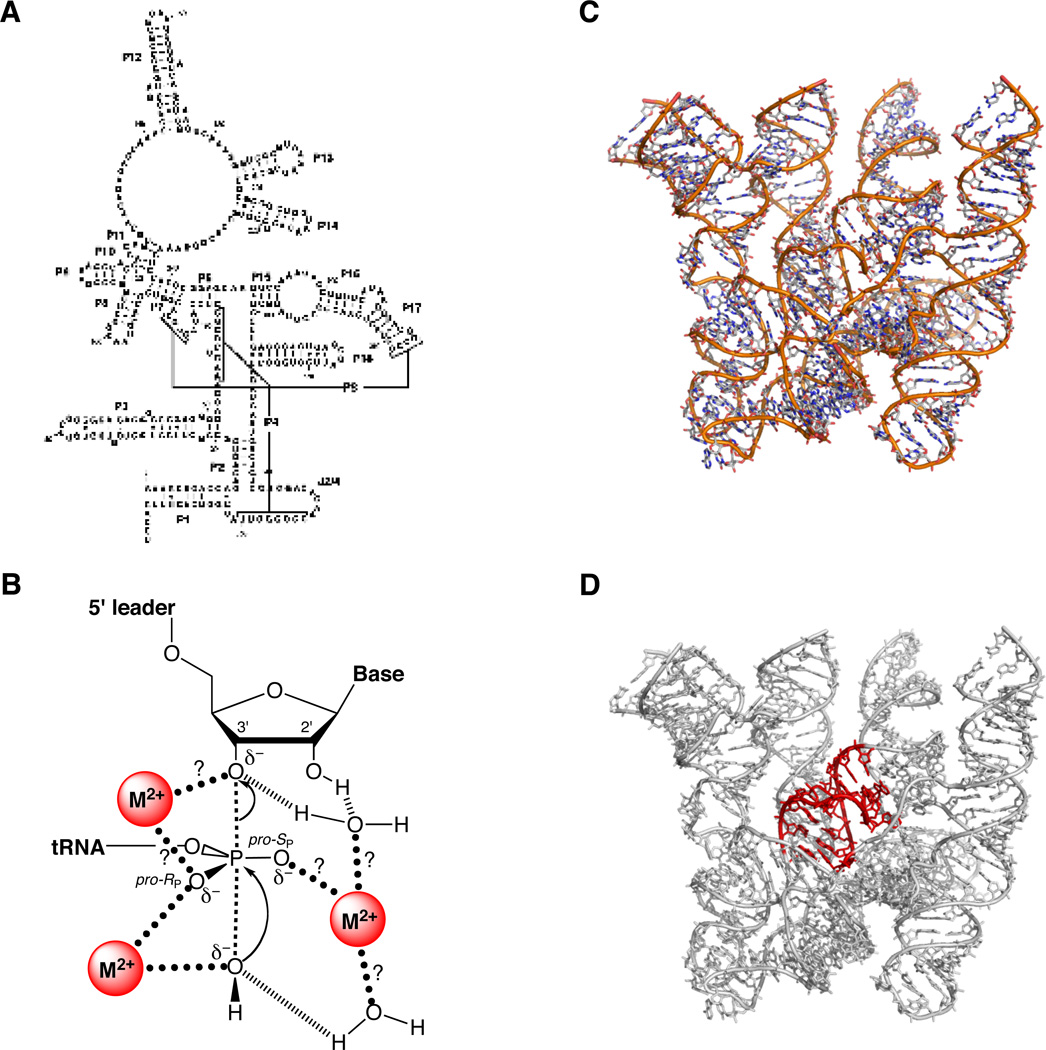
The RNA component of RNase P. (A) Secondary structure of E. coli P RNA. (B) Transition state model for the reaction catalyzed by RNase P. Question marks denote putative metal ion-ligand interactions awaiting experimental verification. [Adapted from (175), with permission.] (C) Crystal structure of the P RNA from B. stearothermophilus at 3.3 Å (PDB file 2A64). (D) Location (in red) of the P4 helix within the B. stearothermophilus P RNA.
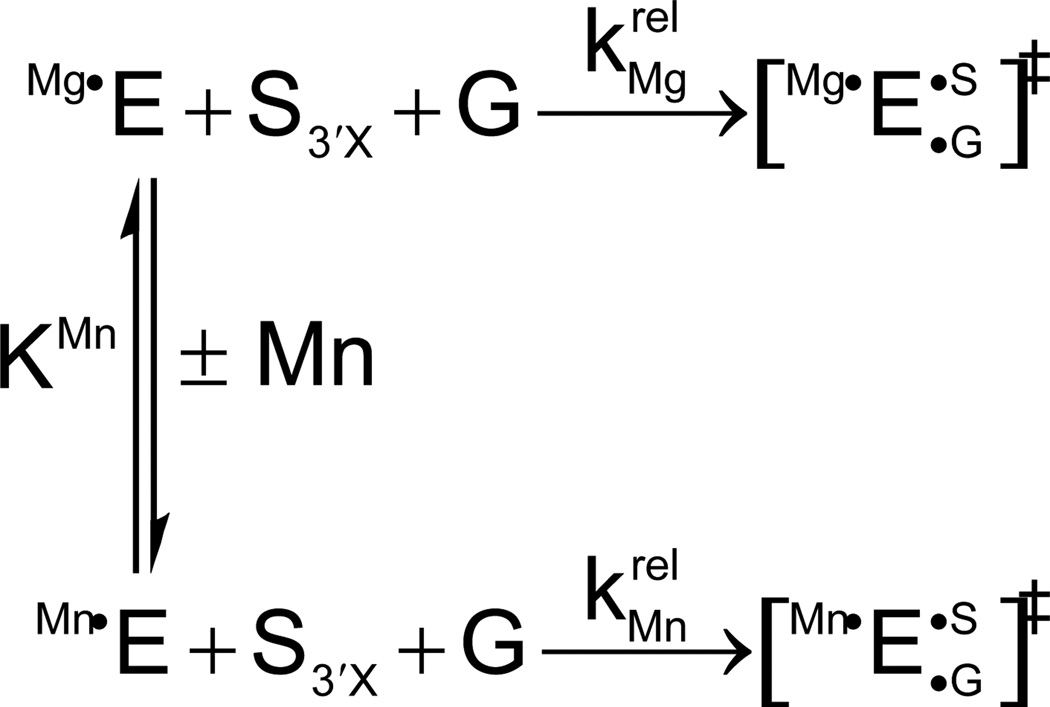
References
-
- Herschlag D, Cech TR. Biochemistry. 1990;29:10159–10171. - PubMed
-
- Pyle AM. Science. 1993;261:709–714. - PubMed
-
- Murray JB, Seyhan AA, Walter NG, Burke JM, Scott WG. Chem Biol. 1998;5:587–595. - PubMed
-
- Nakano S, Chadalavada DM, Bevilacqua PC. Science. 2000;287:1493–1497. - PubMed
-
- Perrotta AT, Been MD. Biochemistry. 2006;45:11357–11365. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
