

Scaffolds: interaction platforms for cellular signalling circuits
- PMID: 19651513
- PMCID: PMC3073007
- DOI: 10.1016/j.tcb.2009.05.007
Scaffolds: interaction platforms for cellular signalling circuits
Abstract
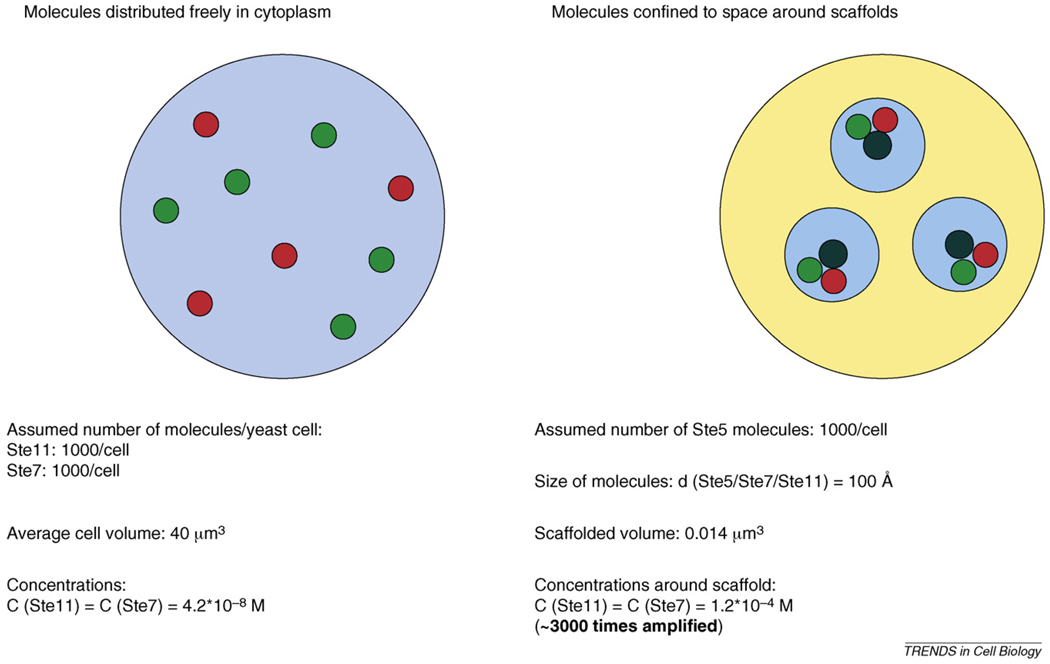
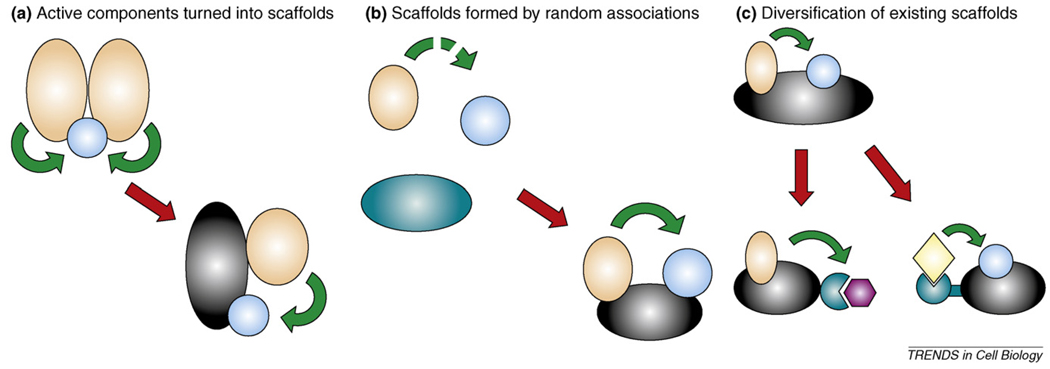
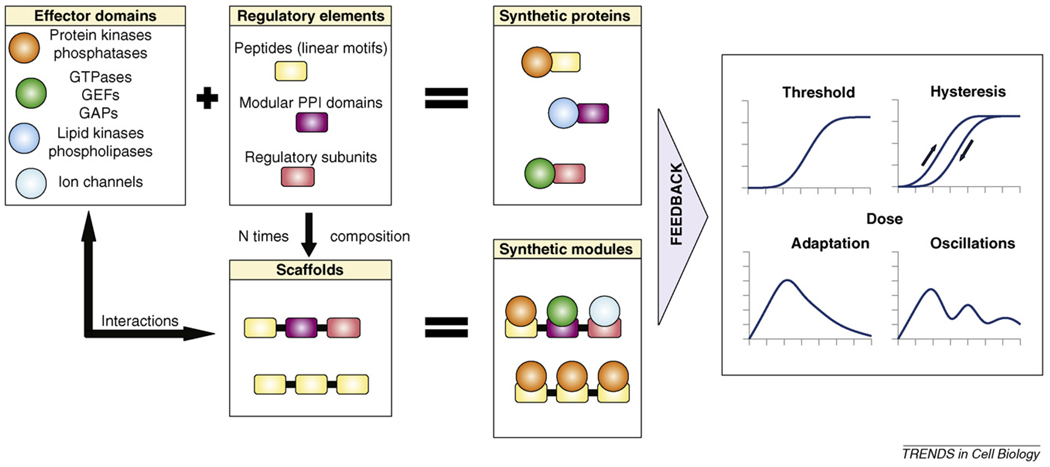
Scaffold proteins influence cellular signalling by binding to multiple signalling enzymes, receptors or ion channels. Although normally devoid of catalytic activity, they have a big impact on controlling the flow of signalling information. By assembling signalling proteins into complexes, they play the part of signal processing hubs. As we learn more about the way signalling components are linked into natural signalling circuits, researchers are becoming interested in building non-natural signalling pathways to test our knowledge and/or to intentionally reprogram cellular behaviour. In this review, we discuss the role of scaffold proteins as efficient tools for assembling intracellular signalling complexes, both natural and artificial.
Figures
Comment in
-
Finding scaffold proteins in interactomes.Trends Cell Biol. 2010 Jan;20(1):2-4. doi: 10.1016/j.tcb.2009.11.003. Epub 2009 Dec 11. Trends Cell Biol. 2010. PMID: 20005715 No abstract available.
References
-
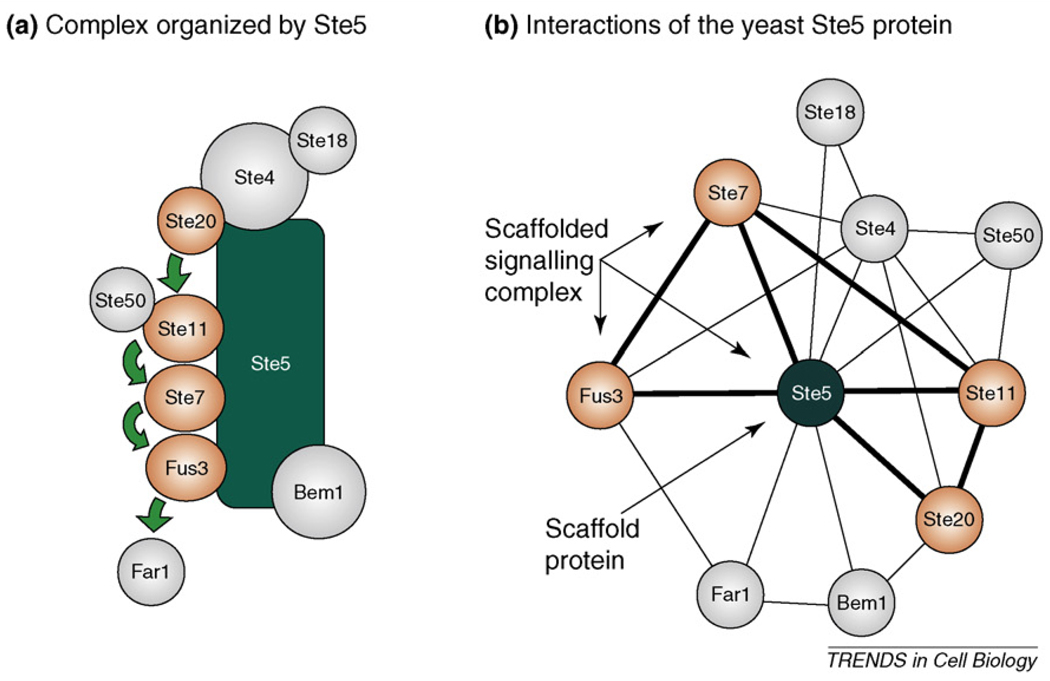
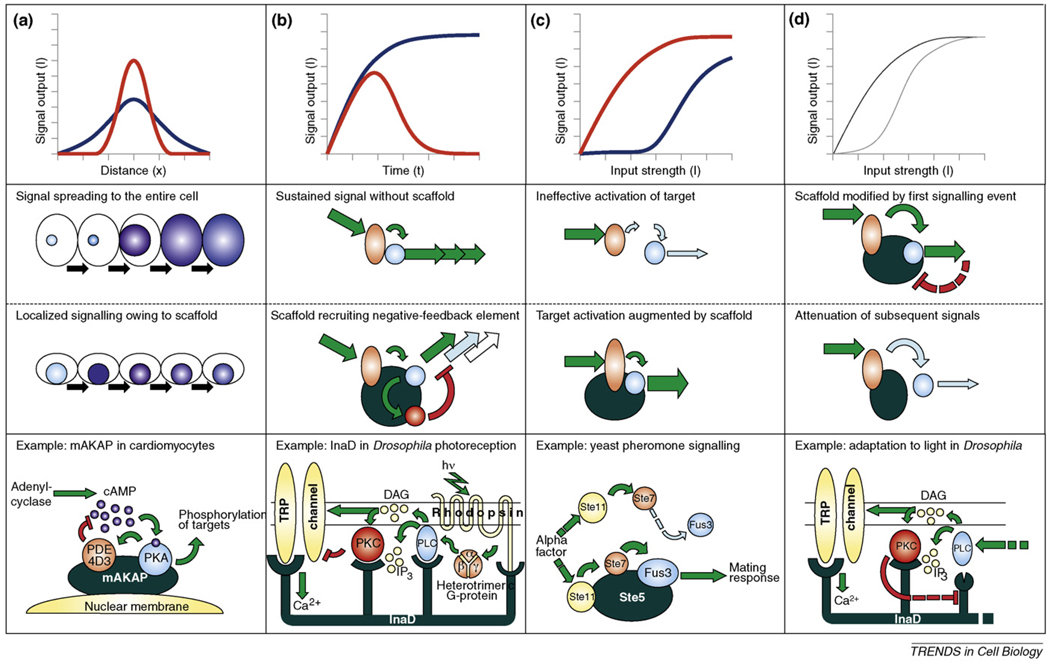
- Choi KY, et al. Ste5 tethers multiple protein kinases in the MAP kinase cascade required for mating in S. cerevisiae. Cell. 1994;78:499–512. - PubMed
-
- Friedman A, Perrimon N. Genetic screening for signal transduction in the era of network biology. Cell. 2007;128:225–231. - PubMed
-
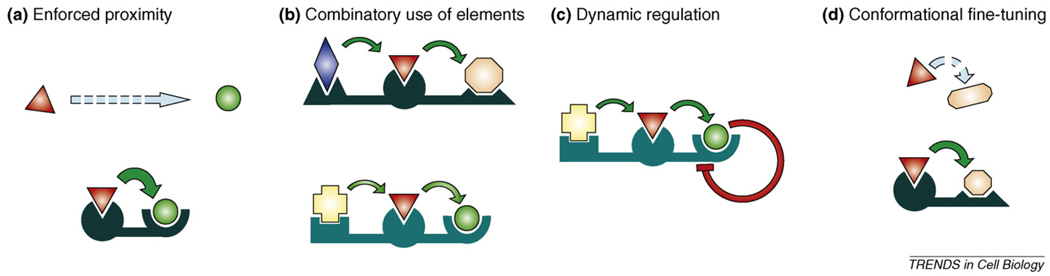
- Ferrrell JE, Jr, Cimprich KA. Enforced proximity in the function of a famous scaffold. Mol. Cell. 2003;11:289–291. - PubMed
-
- Bhattacharyya RP, et al. Domains, motifs, and scaffolds: the role of modular interactions in the evolution and wiring of cell signalling circuits. Annu. Rev. Biochem. 2006;75:655–680. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
