

Structural basis for a human glycosylation disorder caused by mutation of the COG4 gene
- PMID: 19651599
- PMCID: PMC2716380
- DOI: 10.1073/pnas.0901966106
Structural basis for a human glycosylation disorder caused by mutation of the COG4 gene
Abstract
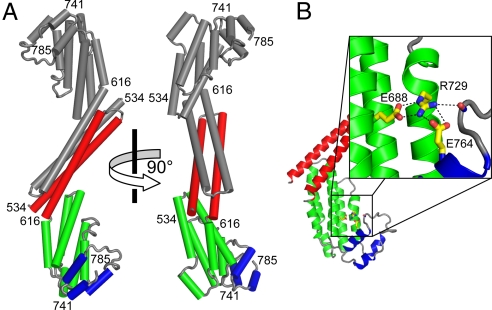
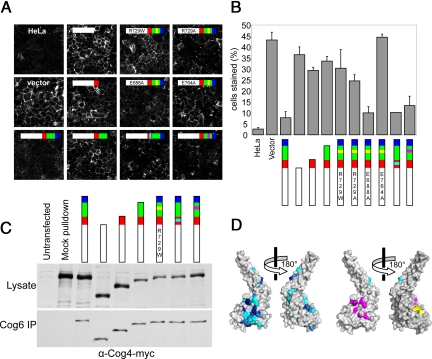
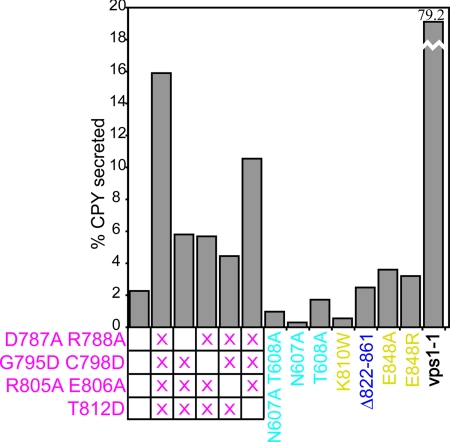
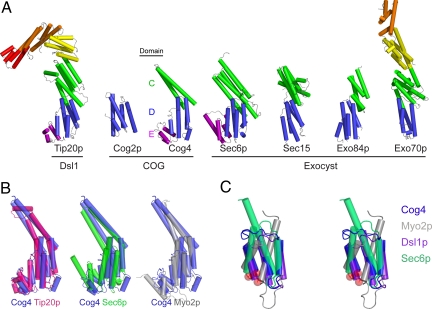
The proper glycosylation of proteins trafficking through the Golgi apparatus depends upon the conserved oligomeric Golgi (COG) complex. Defects in COG can cause fatal congenital disorders of glycosylation (CDGs) in humans. The recent discovery of a form of CDG, caused in part by a COG4 missense mutation changing Arg 729 to Trp, prompted us to determine the 1.9 A crystal structure of a Cog4 C-terminal fragment. Arg 729 is found to occupy a key position at the center of a salt bridge network, thereby stabilizing Cog4's small C-terminal domain. Studies in HeLa cells reveal that this C-terminal domain, while not needed for the incorporation of Cog4 into COG complexes, is essential for the proper glycosylation of cell surface proteins. We also find that Cog4 bears a strong structural resemblance to exocyst and Dsl1p complex subunits. These complexes and others have been proposed to function by mediating the initial tethering between transport vesicles and their membrane targets; the emerging structural similarities provide strong evidence of a common evolutionary origin and may reflect shared mechanisms of action.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Lupashin V, Sztul E. Golgi tethering factors. Biochim Biophys Acta. 2005;1744:325–339. - PubMed
-
- Ungar D, Oka T, Krieger M, Hughson FM. Retrograde transport on the COG railway. Trends Cell Biol. 2006;16:113–120. - PubMed
-
- Walter DM, Paul KS, Waters MG. Purification and characterization of a novel 13 S hetero-oligomeric protein complex that stimulates in vitro Golgi transport. J Biol Chem. 1998;273:29565–29576. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
