

Differential regulation of the central neural cardiorespiratory system by metabotropic neurotransmitters
- PMID: 19651655
- PMCID: PMC2865124
- DOI: 10.1098/rstb.2009.0092
Differential regulation of the central neural cardiorespiratory system by metabotropic neurotransmitters
Abstract
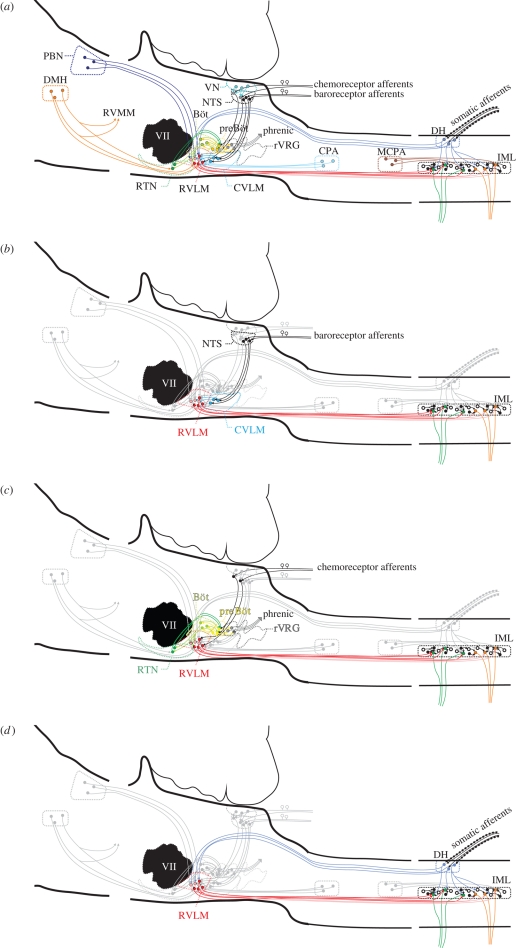
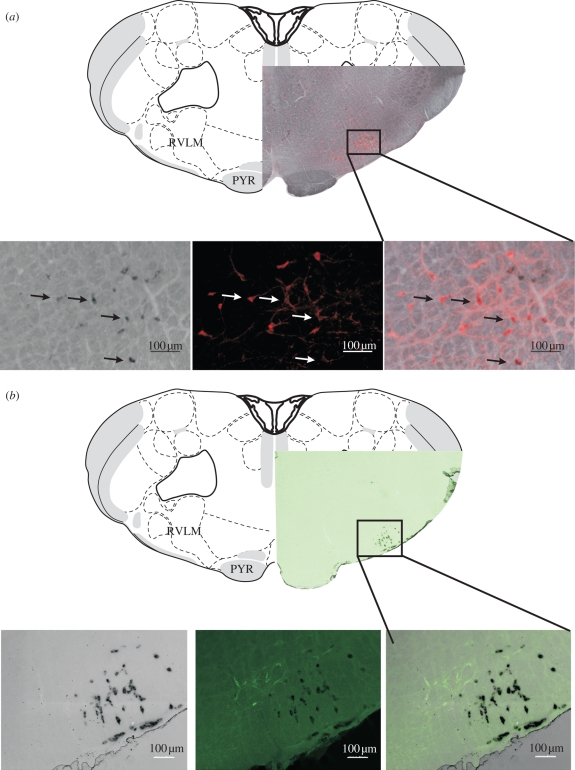
Central neurons in the brainstem and spinal cord are essential for the maintenance of sympathetic tone, the integration of responses to the activation of reflexes and central commands, and the generation of an appropriate respiratory motor output. Here, we will discuss work that aims to understand the role that metabotropic neurotransmitter systems play in central cardiorespiratory mechanisms. It is well known that blockade of glutamatergic, gamma-aminobutyric acidergic and glycinergic pathways causes major or even complete disruption of cardiorespiratory systems, whereas antagonism of other neurotransmitter systems barely affects circulation or ventilation. Despite the lack of an 'all-or-none' role for metabotropic neurotransmitters, they are nevertheless significant in modulating the effects of central command and peripheral adaptive reflexes. Finally, we propose that a likely explanation for the plethora of neurotransmitters and their receptors on cardiorespiratory neurons is to enable differential regulation of outputs in response to reflex inputs, while at the same time maintaining a tonic level of sympathetic activity that supports those organs that significantly autoregulate their blood supply, such as the heart, brain, retina and kidney. Such an explanation of the data now available enables the generation of many new testable hypotheses.
Figures
References
-
- Achour L., Labbe-Jullie C., Scott M. G. H., Marullo S.2008An escort for GPCRs: implications for regulation of receptor density at the cell surface. Trends Pharmacol. Sci. 29, 528–535 (doi:10.1016/j.tips.2008.07.009) - DOI - PubMed
-
- Agnati L. F., Fuxe K., Zoli M., Zini I., Härfstrand A., Toffano G., Goldstein M.1988Morphometrical and microdensitometrical studies on phenylethanolamine-N-methyltransferase- and neuropeptide Y-immunoreactive neurons in the rostral medulla oblongata of the adult and old male rat. Neuroscience 26, 461–478 (doi:10.1016/0306-4522(88)90162-5) - DOI - PubMed
-
- Aicher S. A., Schreihofer A. M., Kraus J. A., Sharma S., Milner T. A., Guyenet P. G.2001Mu-opioid receptors are present in functionally identified sympathoexcitatory neurons in the rat rostral ventrolateral medulla. J. Comp. Neurol. 433, 34–47 (doi:10.1002/cne.1123) - DOI - PubMed
-
- Aicher S. A., Sharma S., Mitchell J. L.2002Co-localization of AMPA receptor subunits in the nucleus of the solitary tract in the rat. Brain Res. 958, 454–458 (doi:10.1016/S0006-8993(02)03693-4) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources