

Intercellular communication of cellular stress monitored by gamma-H2AX induction
- PMID: 19651821
- PMCID: PMC2757548
- DOI: 10.1093/carcin/bgp192
Intercellular communication of cellular stress monitored by gamma-H2AX induction
Abstract
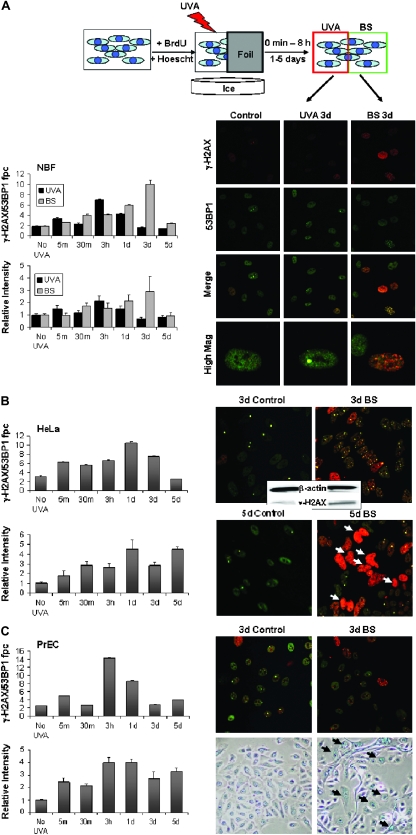
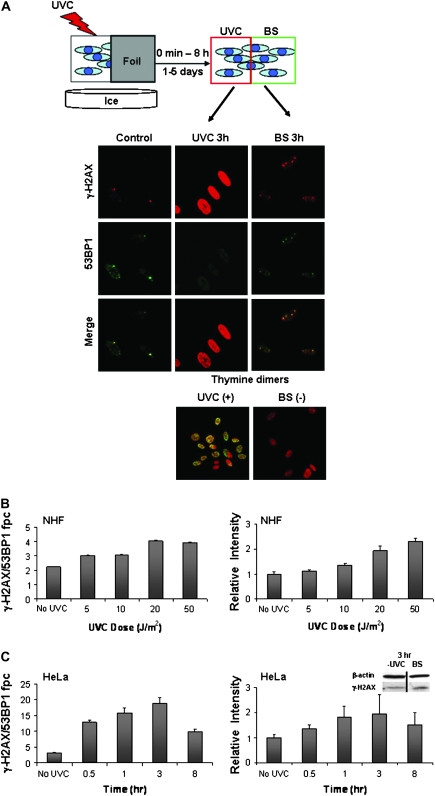
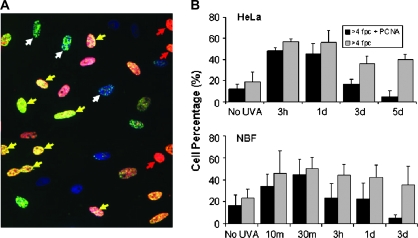
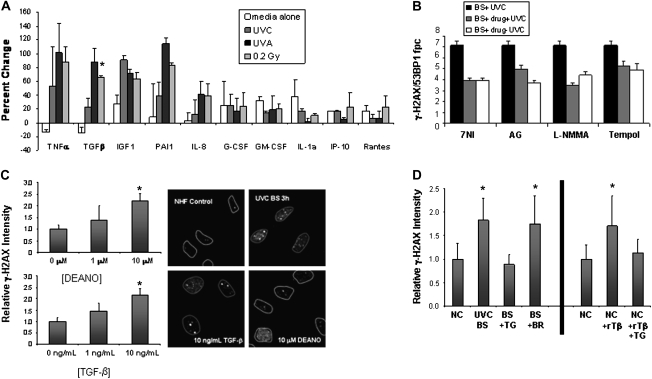
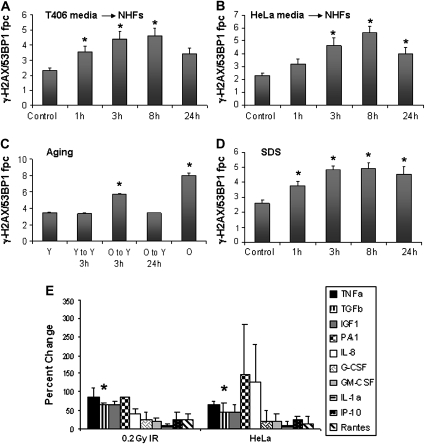
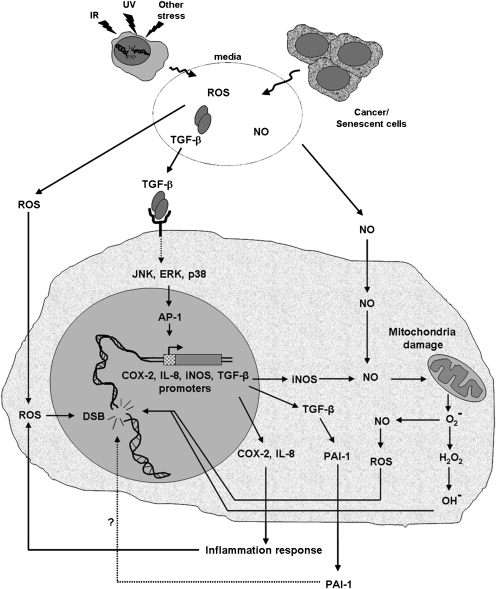
When cells are exposed to ionizing radiation (IR), unexposed cells that share media with damaged cells exhibit similar effects to irradiated cells including increased levels of DNA double-strand breaks (DSBs). Hypothesizing that this effect, known as the radiation-induced bystander effect, may be a specific instance of communication between damaged and undamaged cells regardless of damage source, we demonstrated that exposure of target cells to non-IR induces bystander damage in non-targeted cells as measured by gamma-H2AX and 53BP1 focal formation. Initially, bystander damage was found primarily in S-phase cells, but at later times, non-S-phase cells were also affected. In addition, media from undamaged malignant and senescent cells also was found to induce DSBs in primary cultures. Media conditioned on cells targeted with either ionizing or non-IR as well as on undamaged malignant and senescent cells contained elevated levels of several cytokines. One of these, transforming growth factor beta (TGF-beta), and nitric oxide (NO) were found to elevate numbers of gamma-H2AX/53BP1 foci in normal cell cultures similar to levels found in bystander cells, and this elevation was abrogated by NO synthase inhibitors, TGF-beta blocking antibody and antioxidants. These findings support the hypothesis that damage in bystander cells results from their exposure to cytokines or reactive compounds released from stressed cells, regardless of damage source. These results have implications for oncogenesis in that they indicate that damaged normal cells or undamaged tumor cells may induce genomic instability, leading to an increased risk of oncogenic transformation in other cells with which they share media or contact directly.
Figures
References
-
- Nagasawa H, et al. Induction of sister chromatid exchanges by extremely low doses of alpha-particles. Cancer Res. 1992;52:6394–6396. - PubMed
-
- Nagasawa H, et al. Bystander effect for chromosomal aberrations induced in wild-type and repair deficient CHO cells by low fluences of alpha particles. Mutat. Res. 2002;508:121–129. - PubMed
-
- Azzam EI, et al. Oxidative metabolism modulates signal transduction and micronucleus formation in bystander cells from alpha-particle-irradiated normal human fibroblast cultures. Cancer Res. 2002;62:5436–5442. - PubMed
-
- Prise KM, et al. What role for DNA damage and repair in the bystander response? Mutat. Res. 2006;597:1–4. - PubMed
-
- Hu B, et al. The time and spatial effects of bystander response in mammalian cells induced by low dose radiation. Carcinogenesis. 2006;27:245–251. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
