

Rapid evolution of sex pheromone-producing enzyme expression in Drosophila
- PMID: 19652700
- PMCID: PMC2711336
- DOI: 10.1371/journal.pbio.1000168
Rapid evolution of sex pheromone-producing enzyme expression in Drosophila
Abstract
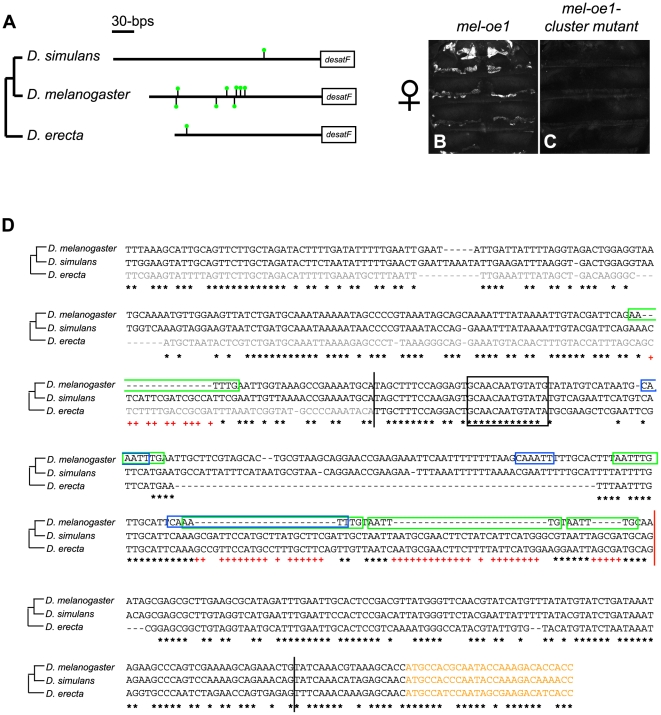
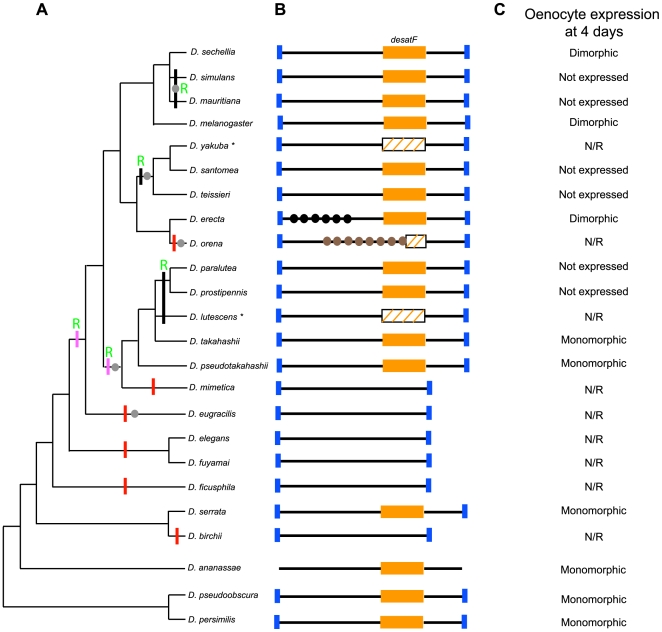
A wide range of organisms use sex pheromones to communicate with each other and to identify appropriate mating partners. While the evolution of chemical communication has been suggested to cause sexual isolation and speciation, the mechanisms that govern evolutionary transitions in sex pheromone production are poorly understood. Here, we decipher the molecular mechanisms underlying the rapid evolution in the expression of a gene involved in sex pheromone production in Drosophilid flies. Long-chain cuticular hydrocarbons (e.g., dienes) are produced female-specifically, notably via the activity of the desaturase DESAT-F, and are potent pheromones for male courtship behavior in Drosophila melanogaster. We show that across the genus Drosophila, the expression of this enzyme is correlated with long-chain diene production and has undergone an extraordinary number of evolutionary transitions, including six independent gene inactivations, three losses of expression without gene loss, and two transitions in sex-specificity. Furthermore, we show that evolutionary transitions from monomorphism to dimorphism (and its reversion) in desatF expression involved the gain (and the inactivation) of a binding-site for the sex-determination transcription factor, DOUBLESEX. In addition, we documented a surprising example of the gain of particular cis-regulatory motifs of the desatF locus via a set of small deletions. Together, our results suggest that frequent changes in the expression of pheromone-producing enzymes underlie evolutionary transitions in chemical communication, and reflect changing regimes of sexual selection, which may have contributed to speciation among Drosophila.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

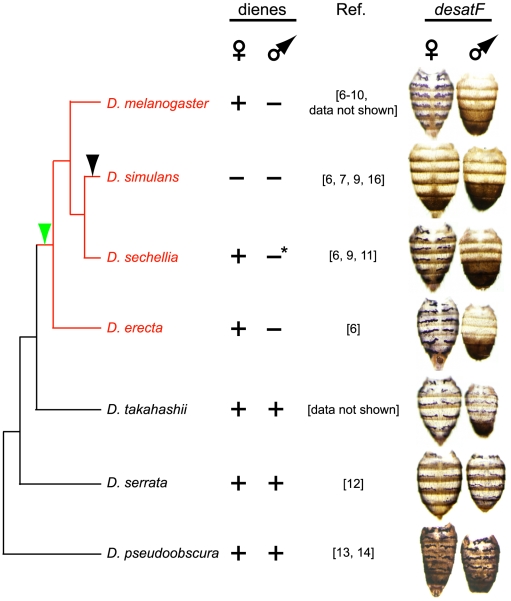
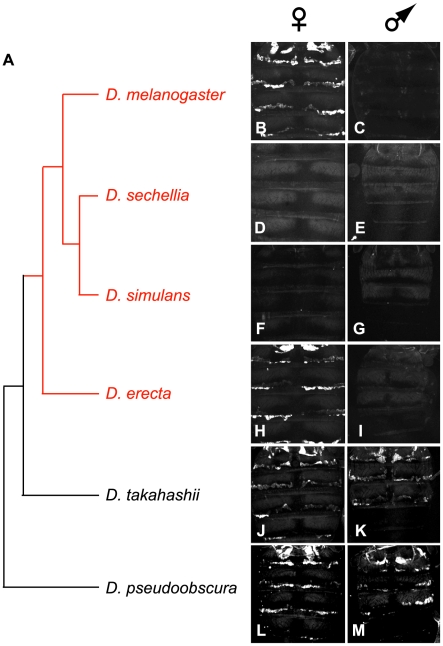
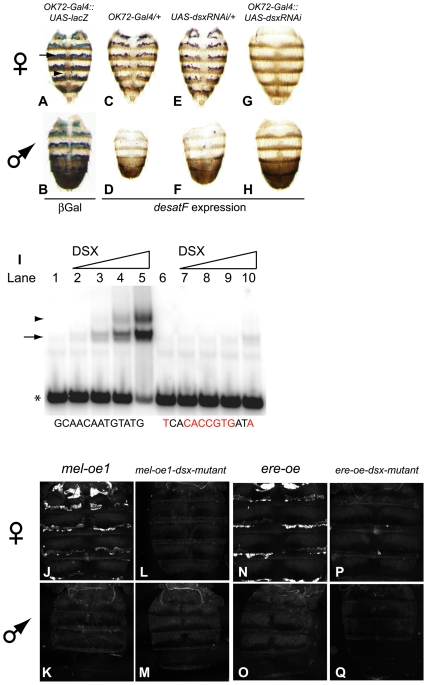
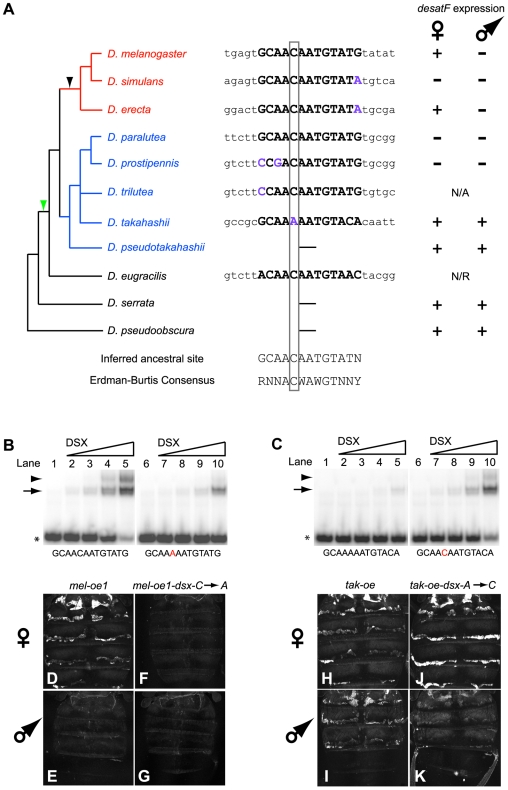
References
-
- Wyatt T. D. Pheromones and animal behaviour: communication by smell and taste. Cambridge; New York: Cambridge University Press; 2003. p. xv.
-
- Johansson B. G, Jones T. M. The role of chemical communication in mate choice. Biol Rev Camb Philos Soc. 2007;82:265–289. - PubMed
-
- Smadja C, Butlin R. K. On the scent of speciation: the chemosensory system and its role in premating isolation. Heredity. 2009;102:77–97. - PubMed
-
- Ellegren H, Parsch J. The evolution of sex-biased genes and sex-biased gene expression. Nat Rev Genet. 2007;8:689–698. - PubMed
-
- Ferveur J. F. Cuticular hydrocarbons: their evolution and roles in Drosophila pheromonal communication. Behav Genet. 2005;35:279–295. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
