

AftD, a novel essential arabinofuranosyltransferase from mycobacteria
- PMID: 19654261
- PMCID: PMC2757576
- DOI: 10.1093/glycob/cwp116
AftD, a novel essential arabinofuranosyltransferase from mycobacteria
Abstract
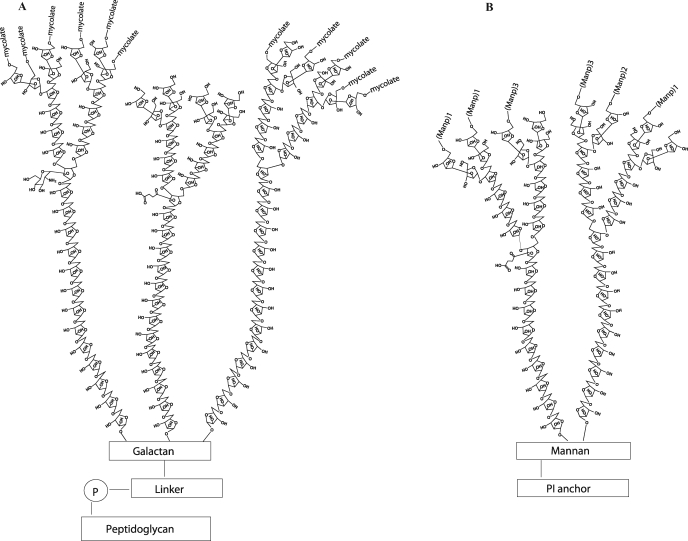
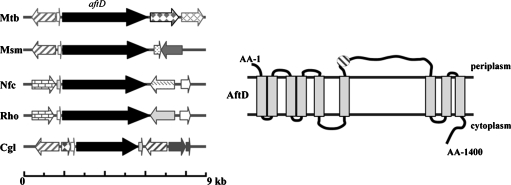
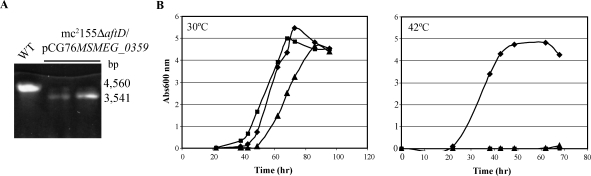
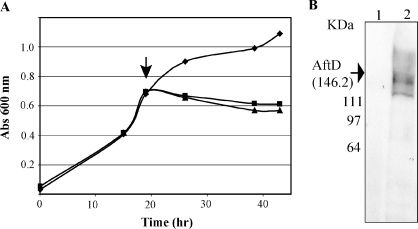
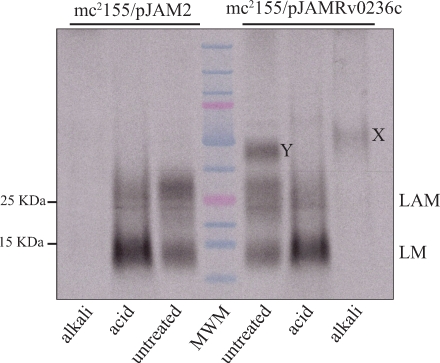
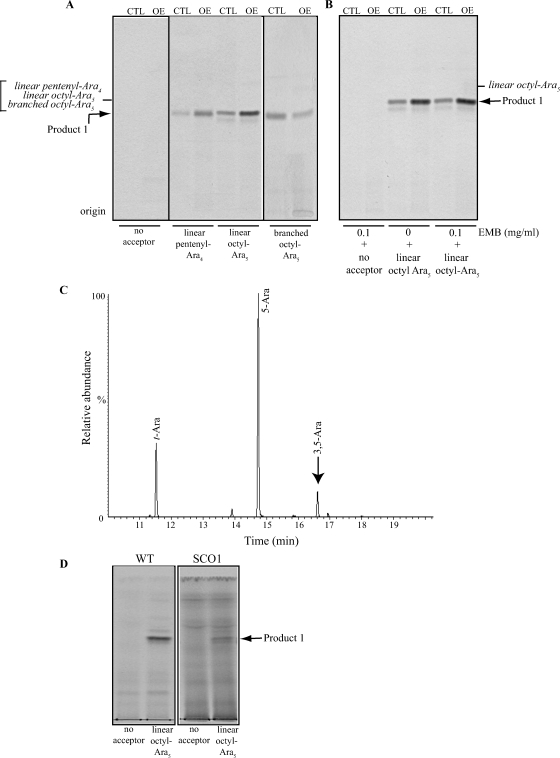
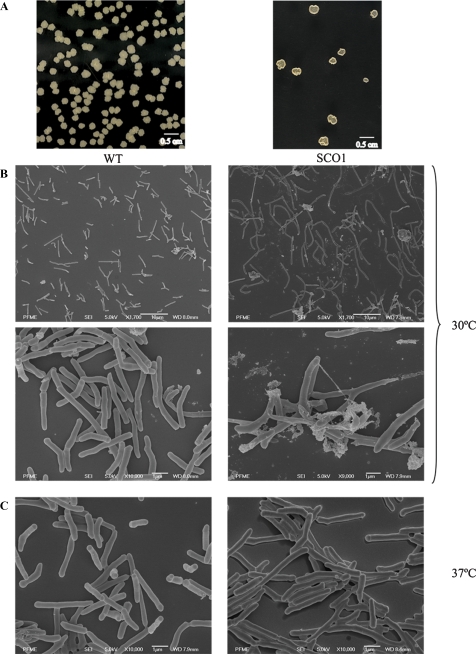
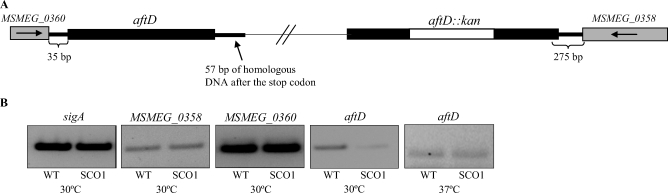
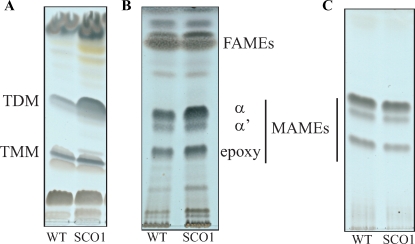
Arabinogalactan (AG) and lipoarabinomannan (LAM) are the two major cell wall (lipo)polysaccharides of mycobacteria. They share arabinan chains made of linear segments of alpha-1,5-linked D-Araf residues with some alpha-1,3-branching, the biosynthesis of which offers opportunities for new chemotherapeutics. In search of the missing arabinofuranosyltransferases (AraTs) responsible for the formation of the arabinan domains of AG and LAM in Mycobacterium tuberculosis, we identified Rv0236c (AftD) as a putative membrane-associated polyprenyl-dependent glycosyltransferase. AftD is 1400 amino acid-long, making it the largest predicted glycosyltransferase of its class in the M. tuberculosis genome. Assays using cell-free extracts from recombinant Mycobacterium smegmatis and Corynebacterium glutamicum strains expressing different levels of aftD indicated that this gene encodes a functional AraT with alpha-1,3-branching activity on linear alpha-1,5-linked neoglycolipid acceptors in vitro. The disruption of aftD in M. smegmatis resulted in cell death and a decrease in its activity caused defects in cell division, reduced growth, alteration of colonial morphology, and accumulation of trehalose dimycolates in the cell envelope. Overexpression of aftD in M. smegmatis, in contrast, induced the accumulation of two arabinosylated compounds with carbohydrate backbones reminiscent of that of LAM and a degree of arabinosylation dependent on aftD expression levels. Altogether, our results thus indicate that AftD is an essential AraT involved in the synthesis of the arabinan domain of major mycobacterial cell envelope (lipo)polysaccharides.
Figures
References
-
- Alderwick LJ, Seidel M, Sahm H, Besra GS, Eggeling L. Identification of a novel arabinosyl transferase (AftA) involved in cell wall arabinan biosynthesis in Mycobacterium tuberculosis. J Biol Chem. 2006;281:15653–15661. - PubMed
-
- Berg S, Kaur D, Jackson M, Brennan PJ. The glycosyltransferases of Mycobacterium tuberculosis- roles in the synthesis of arabinogalactan, lipoarabinomannan, and other glycoconjugates. Glycobiology. 2007;17:35R–56R. - PubMed
-
- Berg S, Starbuck J, Torrelles JB, Vissa VD, Crick DC, Chatterjee C, Brennan PJ. Roles of the conserved proline and glycosyltransferase motifs of EmbC in biosynthesis of lipoarabinomannan. J Biol Chem. 2005;280:5651–5663. - PubMed
-
- Bhamidi S, Scherman MS, Rithner CD, Prenni JE, Chatterjee D, Khoo K-H, McNeil MR. The identification and location of succinyl residues and the characterization of the interior arabinan region allows for a model of the complete primary structure of Mycobacterium tuberculosis mycolyl arabinogalactan. J Biol Chem. 2008;283:12992–13000. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
