

Genomic analysis of a sexually-selected character: EST sequencing and microarray analysis of eye-antennal imaginal discs in the stalk-eyed fly Teleopsis dalmanni (Diopsidae)
- PMID: 19656405
- PMCID: PMC2737001
- DOI: 10.1186/1471-2164-10-361
Genomic analysis of a sexually-selected character: EST sequencing and microarray analysis of eye-antennal imaginal discs in the stalk-eyed fly Teleopsis dalmanni (Diopsidae)
Abstract
Background: Many species of stalk-eyed flies (Diopsidae) possess highly-exaggerated, sexually dimorphic eye-stalks that play an important role in the mating system of these flies. Eye-stalks are increasingly being used as a model system for studying sexual selection, but little is known about the genetic mechanisms producing variation in these ornamental traits. Therefore, we constructed an EST database of genes expressed in the developing eye-antennal imaginal disc of the highly dimorphic species Teleopsis dalmanni. We used this set of genes to construct microarray slides and compare patterns of gene expression between lines of flies with divergent eyespan.
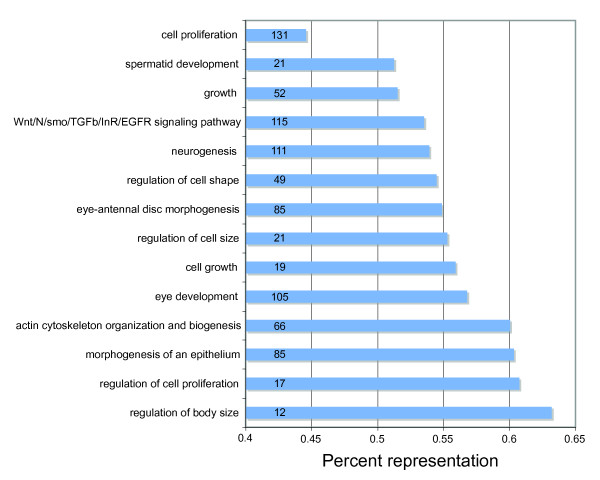
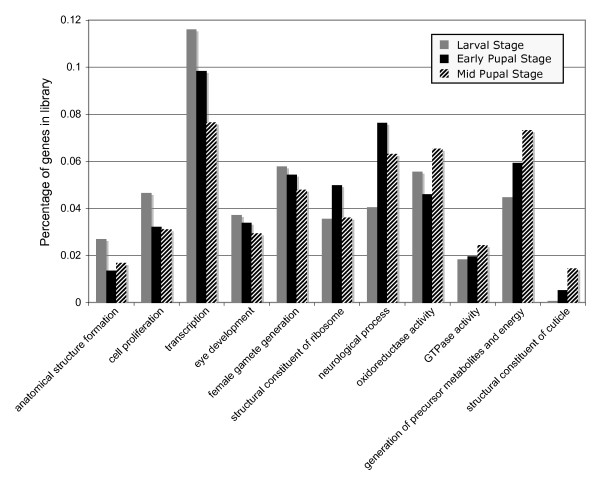
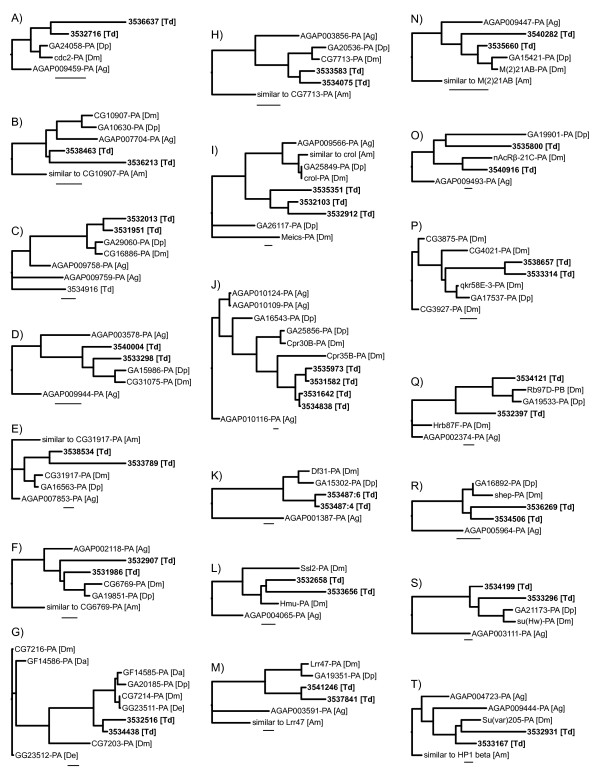
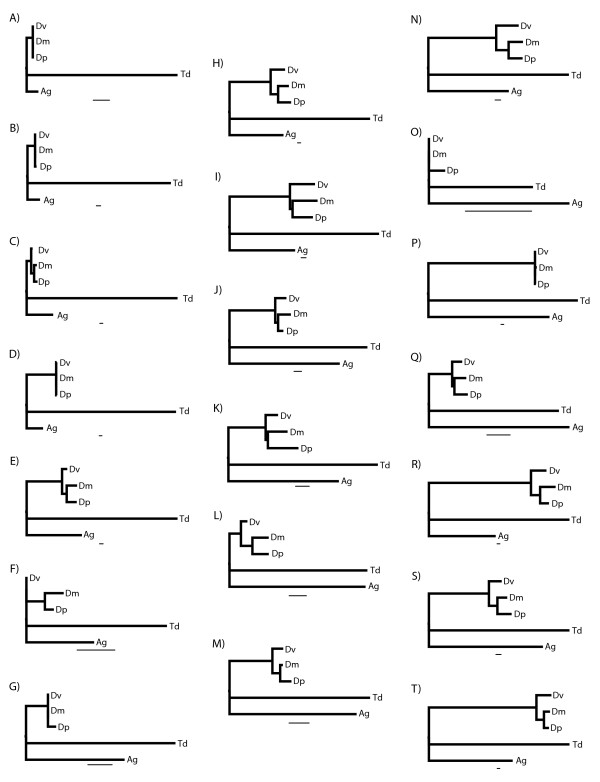
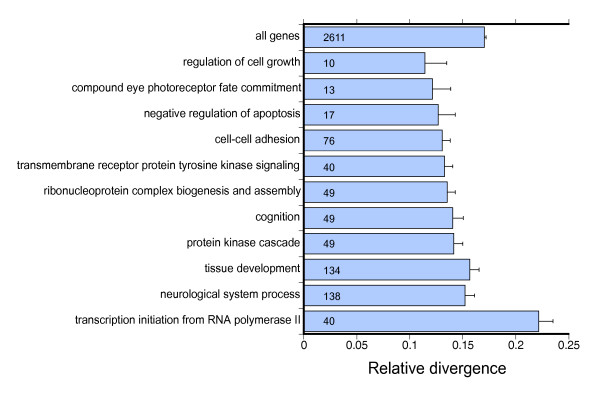
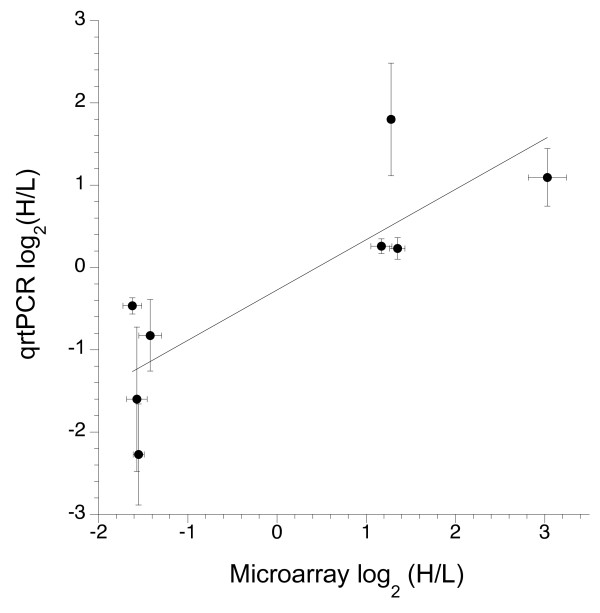
Results: We generated 33,229 high-quality ESTs from three non-normalized libraries made from the developing eye-stalk tissue at different developmental stages. EST assembly and annotation produced a total of 7,066 clusters comprising 3,424 unique genes with significant sequence similarity to a protein in either Drosophila melanogaster or Anopheles gambiae. Comparisons of the transcript profiles at different stages reveal a developmental shift in relative expression from genes involved in anatomical structure formation, transcription, and cell proliferation at the larval stage to genes involved in neurological processes and cuticle production during the pupal stages. Based on alignments of the EST fragments to homologous sequences in Drosophila and Anopheles, we identified 20 putative gene duplication events in T. dalmanni and numerous genes undergoing significantly faster rates of evolution in T. dalmanni relative to the other Dipteran species. Microarray experiments identified over 350 genes with significant differential expression between flies from lines selected for high and low relative eyespan but did not reveal any primary biological process or pathway that is driving the expression differences.
Conclusion: The catalogue of genes identified in the EST database provides a valuable framework for a comprehensive examination of the genetic basis of eye-stalk variation. Several candidate genes, such as crooked legs, cdc2, CG31917 and CG11577, emerge from the analysis of gene duplication, protein evolution and microarray gene expression. Additional comparisons of expression profiles between, for example, males and females, and species that differ in eye-stalk sexual dimorphism, are now enabled by these resources.
Figures
References
-
- Pomiankowski A, Møller AP. A resolution of the Lek paradox. Proc R Soc Lond B. 1995;260:21–29. doi: 10.1098/rspb.1995.0054. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
