

Function and assembly of DNA looping, clustering, and microtubule attachment complexes within a eukaryotic kinetochore
- PMID: 19656849
- PMCID: PMC2754927
- DOI: 10.1091/mbc.e09-05-0359
Function and assembly of DNA looping, clustering, and microtubule attachment complexes within a eukaryotic kinetochore
Abstract
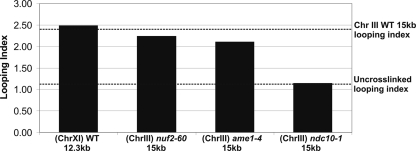
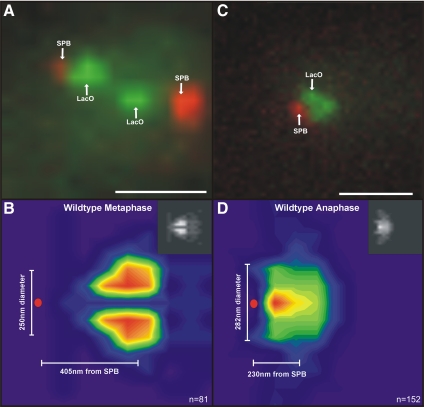
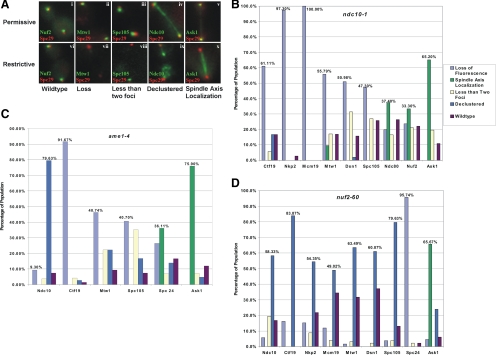
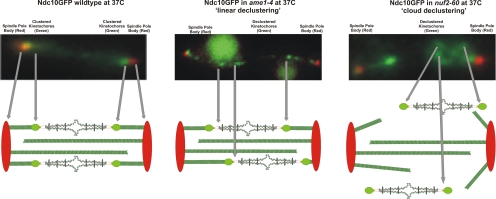
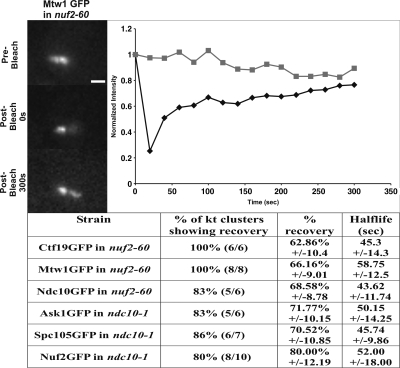
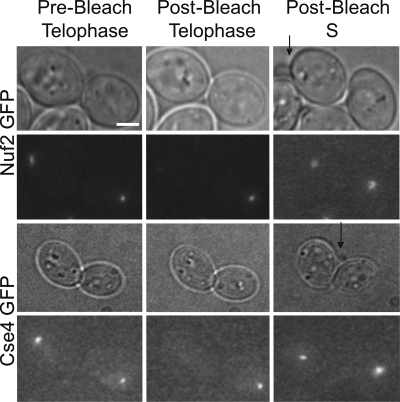
The kinetochore is a complex protein-DNA assembly that provides the mechanical linkage between microtubules and the centromere DNA of each chromosome. Centromere DNA in all eukaryotes is wrapped around a unique nucleosome that contains the histone H3 variant CENP-A (Cse4p in Saccharomyces cerevisiae). Here, we report that the inner kinetochore complex (CBF3) is required for pericentric DNA looping at the Cse4p-containing nucleosome. DNA within the pericentric loop occupies a spatially confined area that is radially displaced from the interpolar central spindle. Microtubule-binding kinetochore complexes are not involved in pericentric DNA looping but are required for the geometric organization of DNA loops around the spindle microtubules in metaphase. Thus, the mitotic segregation apparatus is a composite structure composed of kinetochore and interpolar microtubules, the kinetochore, and organized pericentric DNA loops. The linkage of microtubule-binding to centromere DNA-looping complexes positions the pericentric chromatin loops and stabilizes the dynamic properties of individual kinetochore complexes in mitosis.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
