

Multiscale mechanics of fibrin polymer: gel stretching with protein unfolding and loss of water
- PMID: 19661428
- PMCID: PMC2846107
- DOI: 10.1126/science.1172484
Multiscale mechanics of fibrin polymer: gel stretching with protein unfolding and loss of water
Abstract
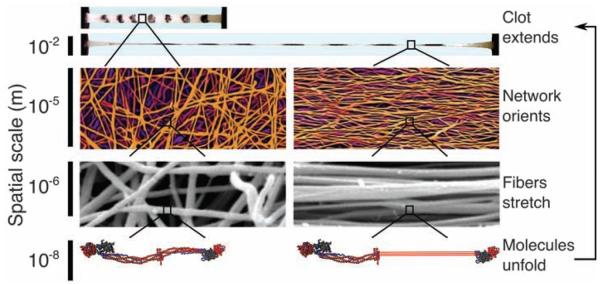
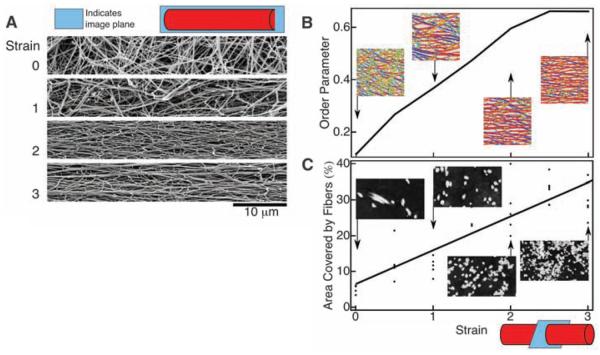
Blood clots and thrombi consist primarily of a mesh of branched fibers made of the protein fibrin. We propose a molecular basis for the marked extensibility and negative compressibility of fibrin gels based on the structural and mechanical properties of clots at the network, fiber, and molecular levels. The force required to stretch a clot initially rises linearly and is accompanied by a dramatic decrease in clot volume and a peak in compressibility. These macroscopic transitions are accompanied by fiber alignment and bundling after forced protein unfolding. Constitutive models are developed to integrate observations at spatial scales that span six orders of magnitude and indicate that gel extensibility and expulsion of water are both manifestations of protein unfolding, which is not apparent in other matrix proteins such as collagen.
Figures


References
-
- Weisel JW. Advances in Protein Chemistry. Vol. 70. Academic Press; London: 2005. pp. 247–299. - PubMed
-
- Laurens N, Koolwijk P, DeMaat MPM. J. Thromb. Haemost. 2006;4:932. - PubMed
-
- Colman RW, et al. Hemostasis and Thrombosis: Basic Principles and Clinical Practice. Lippincott Williams & Wilkins; Philadelphia: 2005.
-
- Weisel JW. Biophys. Chem. 2004;112:267. - PubMed
-
- Collet JP, et al. Arterioscler. Thromb. Vasc. Biol. 2000;20:1354. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
