

Hepatic energy state is regulated by glucagon receptor signaling in mice
- PMID: 19662685
- PMCID: PMC2719934
- DOI: 10.1172/jci38650
Hepatic energy state is regulated by glucagon receptor signaling in mice
Abstract
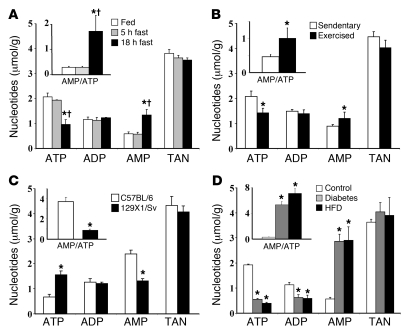
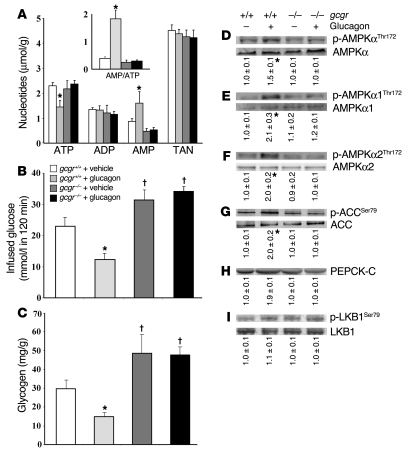
The hepatic energy state, defined by adenine nucleotide levels, couples metabolic pathways with energy requirements. This coupling is fundamental in the adaptive response to many conditions and is impaired in metabolic disease. We have found that the hepatic energy state is substantially reduced following exercise, fasting, and exposure to other metabolic stressors in C57BL/6 mice. Glucagon receptor signaling was hypothesized to mediate this reduction because increased plasma levels of glucagon are characteristic of metabolic stress and because this hormone stimulates energy consumption linked to increased gluconeogenic flux through cytosolic phosphoenolpyruvate carboxykinase (PEPCK-C) and associated pathways. We developed what we believe to be a novel hyperglucagonemic-euglycemic clamp to isolate an increment in glucagon levels while maintaining fasting glucose and insulin. Metabolic stress and a physiological rise in glucagon lowered the hepatic energy state and amplified AMP-activated protein kinase signaling in control mice, but these changes were abolished in glucagon receptor- null mice and mice with liver-specific PEPCK-C deletion. 129X1/Sv mice, which do not mount a glucagon response to hypoglycemia, displayed an increased hepatic energy state compared with C57BL/6 mice in which glucagon was elevated. Taken together, these data demonstrate in vivo that the hepatic energy state is sensitive to glucagon receptor activation and requires PEPCK-C, thus providing new insights into liver metabolism.
Figures





References
-
- Ercan-Fang N., et al. Integrated effects of multiple modulators on human liver glycogen phosphorylase a. Am. J. Physiol. Endocrinol. Metab. 2002;283:E29–E37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
