

Confirmation and fine-mapping of a major QTL for resistance to infectious pancreatic necrosis in Atlantic salmon (Salmo salar): population-level associations between markers and trait
- PMID: 19664221
- PMCID: PMC2728743
- DOI: 10.1186/1471-2164-10-368
Confirmation and fine-mapping of a major QTL for resistance to infectious pancreatic necrosis in Atlantic salmon (Salmo salar): population-level associations between markers and trait
Abstract
Background: Infectious pancreatic necrosis (IPN) is one of the most prevalent and economically devastating diseases in Atlantic salmon (Salmo salar) farming worldwide. The disease causes large mortalities at both the fry- and post-smolt stages. Family selection for increased IPN resistance is performed through the use of controlled challenge tests, where survival rates of sib-groups are recorded. However, since challenge-tested animals cannot be used as breeding candidates, within-family selection is not performed and only half of the genetic variation for IPN resistance is being exploited. DNA markers linked to quantitative trait loci (QTL) affecting IPN resistance would therefore be a powerful selection tool. The aim of this study was to identify and fine-map QTL for IPN-resistance in Atlantic salmon, for use in marker-assisted selection to increase the rate of genetic improvement for this trait.
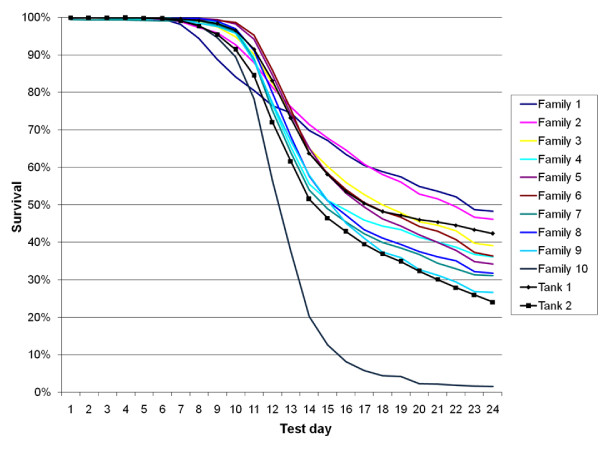
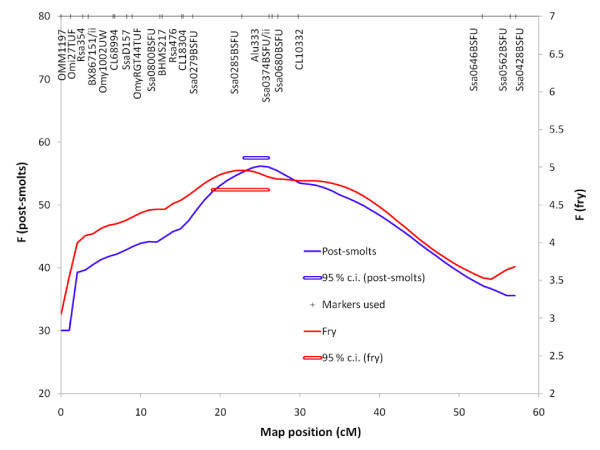
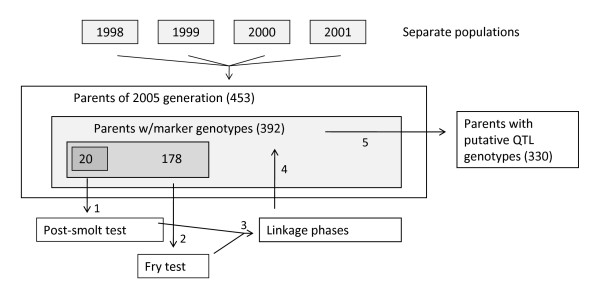
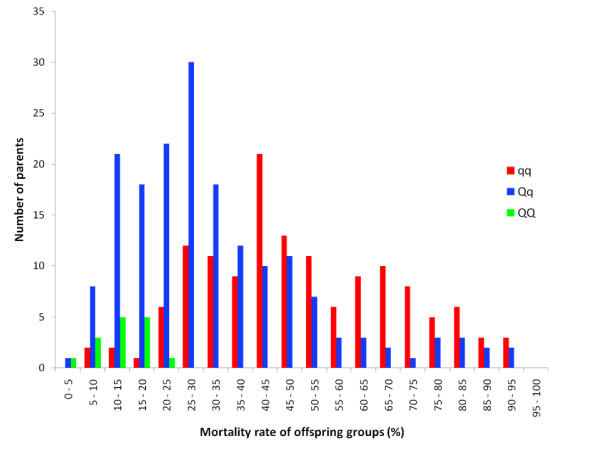
Results: A genome scan was carried out using 10 large full-sib families of challenge-tested Atlantic salmon post-smolts and microsatellite markers distributed across the genome. One major QTL for IPN-resistance was detected, explaining 29% and 83% of the phenotypic and genetic variances, respectively. This QTL mapped to the same location as a QTL recently detected in a Scottish Atlantic salmon population. The QTL was found to be segregating in 10 out of 20 mapping parents, and subsequent fine-mapping with additional markers narrowed the QTL peak to a 4 cM region on linkage group 21. Challenge-tested fry were used to show that the QTL had the same effect on fry as on post-smolt, with the confidence interval for QTL position in fry overlapping the confidence interval found in post-smolts. A total of 178 parents were tested for segregation of the QTL, identifying 72 QTL-heterozygous parents. Genotypes at QTL-heterozygous parents were used to determine linkage phases between alleles at the underlying DNA polymorphism and alleles at single markers or multi-marker haplotypes. One four-marker haplotype was found to be the best predictor of QTL alleles, and was successfully used to deduce genotypes of the underlying polymorphism in 72% of the parents of the next generation within a breeding nucleus. A highly significant population-level correlation was found between deduced alleles at the underlying polymorphism and survival of offspring groups in the fry challenge test, parents with the three deduced genotypes (QQ, Qq, qq) having mean offspring mortality rates of 0.13, 0.32, and 0.49, respectively. The frequency of the high-resistance allele (Q) in the population was estimated to be 0.30. Apart from this major QTL, one other experiment-wise significant QTL for IPN-resistance was detected, located on linkage group 4.
Conclusion: The QTL confirmed in this study represents a case of a major gene explaining the bulk of genetic variation for a presumed complex trait. QTL genotypes were deduced within most parents of the 2005 generation of a major breeding company, providing a solid framework for linkage-based MAS within the whole population in subsequent generations. Since haplotype-trait associations valid at the population level were found, there is also a potential for MAS based on linkage disequilibrium (LD). However, in order to use MAS across many generations without reassessment of linkage phases between markers and the underlying polymorphism, the QTL needs to be positioned with even greater accuracy. This will require higher marker densities than are currently available.
Figures
References
-
- Wedemeyer G. Physiology of fish in intensive culture systems. London: Chapman & Hall; 1996.
-
- Ariel E, Olesen NJ. Finfish in aquaculture and their diseases – a retrospective view in the European Community. Bulletin of the European Association of Fish Pathologists. 2002;22:72–85.
-
- Brun E. Epidemiology. In: Skjelstad B, editor. IPN in salmonids: A review. Oslo: The Fisheriers and Aquaculture Industries Research Fund (FHF); 2003.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
