

Telomere extension occurs at most chromosome ends and is uncoupled from fill-in in human cancer cells
- PMID: 19665970
- PMCID: PMC2726829
- DOI: 10.1016/j.cell.2009.05.026
Telomere extension occurs at most chromosome ends and is uncoupled from fill-in in human cancer cells
Abstract
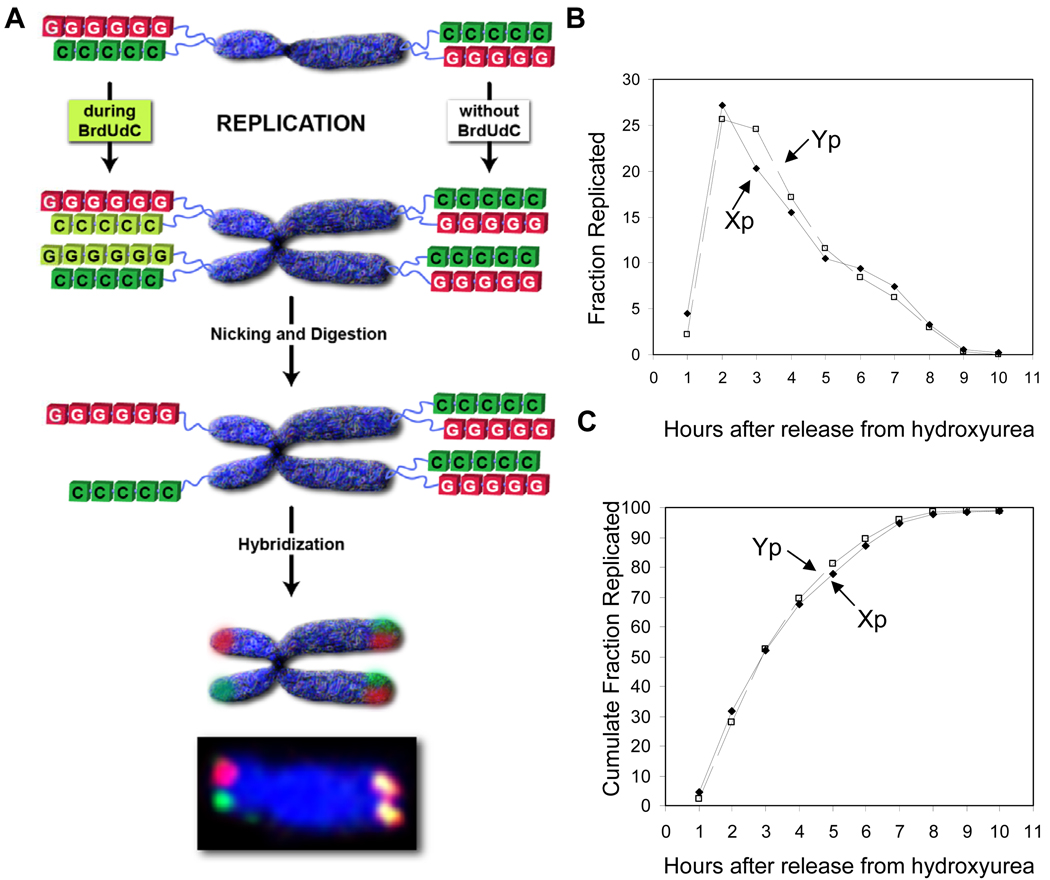
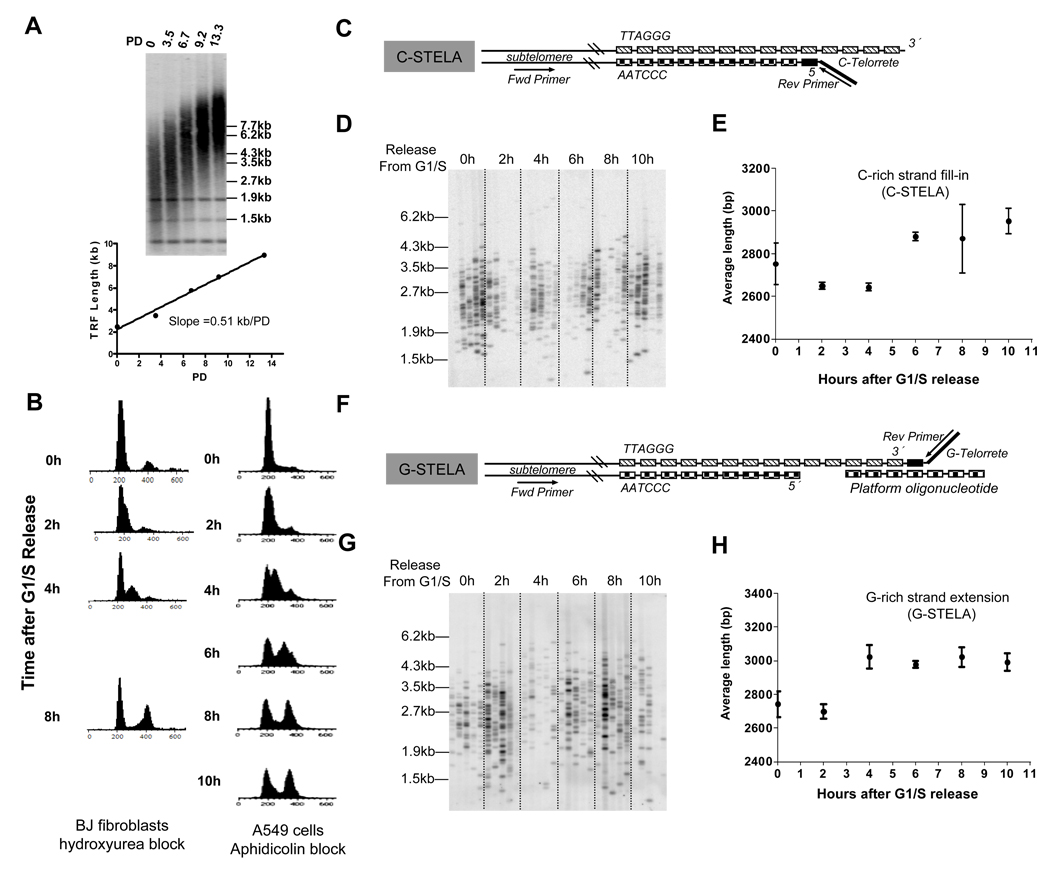
Telomeres are thought to be maintained by the preferential recruitment of telomerase to the shortest telomeres. The extension of the G-rich telomeric strand by telomerase is also believed to be coordinated with the complementary synthesis of the C strand by the conventional replication machinery. However, we show that under telomere length-maintenance conditions in cancer cells, human telomerase extends most chromosome ends during each S phase and is not preferentially recruited to the shortest telomeres. Telomerase rapidly extends the G-rich strand following telomere replication but fill-in of the C strand is delayed into late S phase. This late C-strand fill-in is not executed by conventional Okazaki fragment synthesis but by a mechanism using a series of small incremental steps. These findings highlight differences between telomerase actions during steady state versus nonequilibrium conditions and reveal steps in the human telomere maintenance pathway that may provide additional targets for the development of anti-telomerase therapeutics.
Figures





Comment in
-
Human telomerase caught in the act.Cell. 2009 Aug 7;138(3):432-4. doi: 10.1016/j.cell.2009.07.018. Cell. 2009. PMID: 19665967
References
-
- Bailey SM, Goodwin EH, Cornforth MN. Strand-specific fluorescence in situ hybridization: the CO-FISH family. Cytogenet Genome Res. 2004;107:14–17. - PubMed
-
- Baird DM, Rowson J, Wynford-Thomas D, Kipling D. Extensive allelic variation and ultrashort telomeres in senescent human cells. Nat Genet. 2003;33:203–207. - PubMed
-
- Bianchi A, Shore D. How telomerase reaches its end: mechanism of telomerase regulation by the telomeric complex. Mol Cell. 2008;31:153–165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
