

Structural and functional relationships of the steroid hormone receptors' N-terminal transactivation domain
- PMID: 19666041
- PMCID: PMC3074935
- DOI: 10.1016/j.steroids.2009.07.012
Structural and functional relationships of the steroid hormone receptors' N-terminal transactivation domain
Abstract
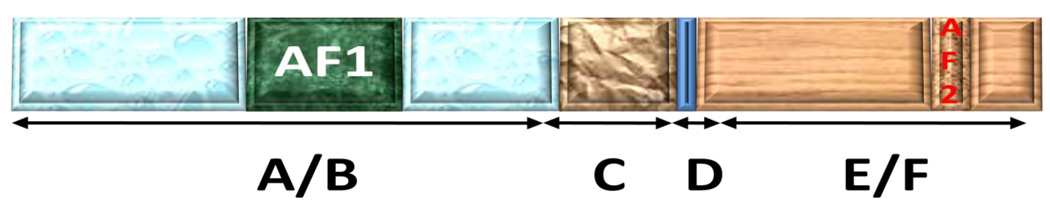
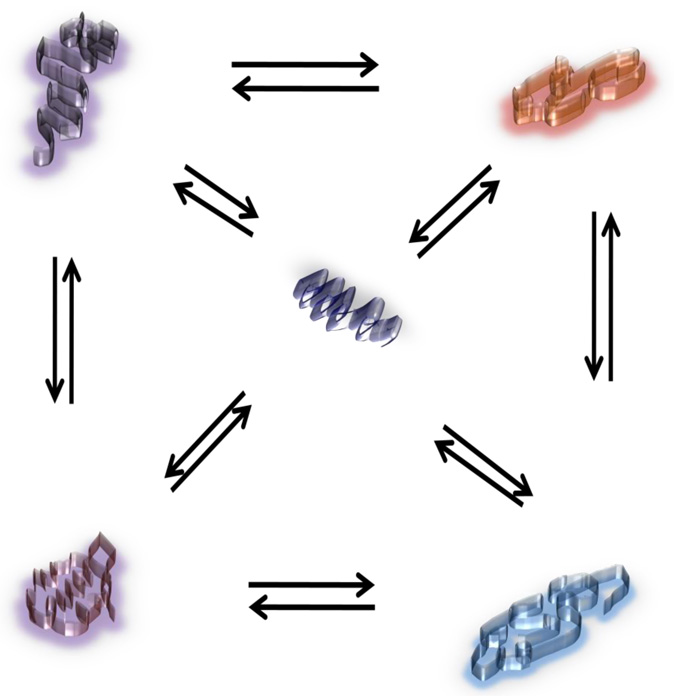
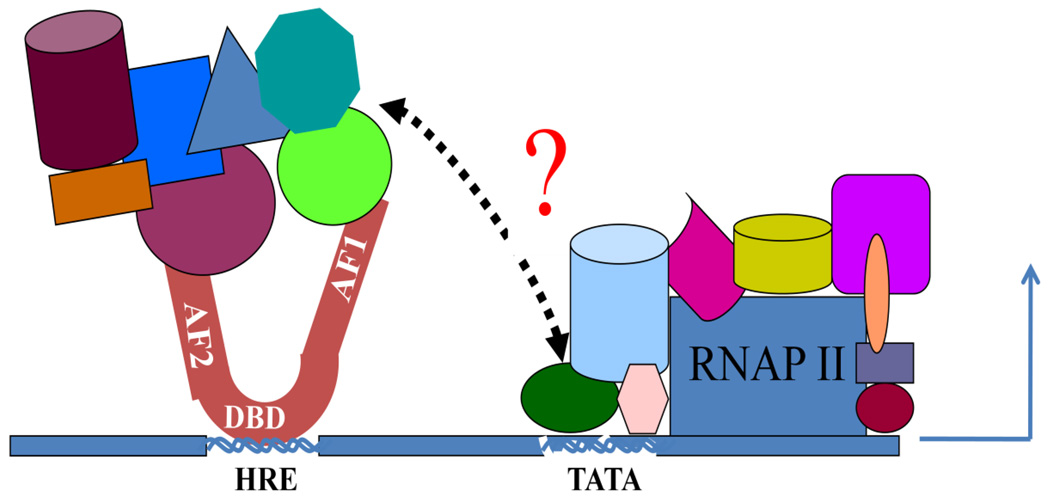
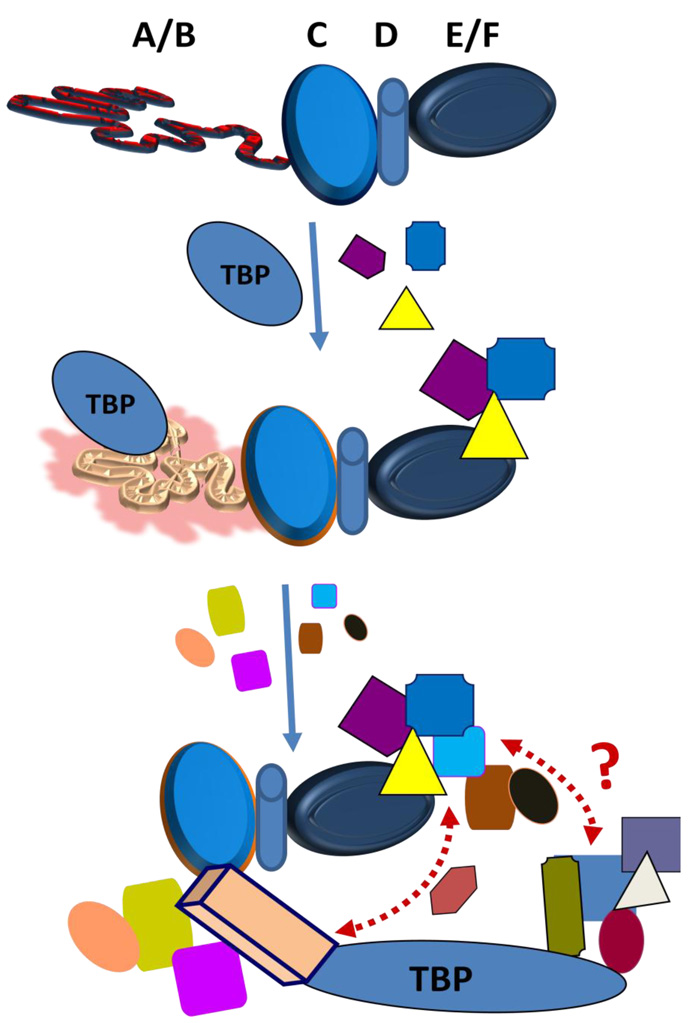
Steroid hormone receptors are members of a family of ligand inducible transcription factors, and regulate the transcriptional activation of target genes by recruiting coregulatory proteins to the pre-initiation machinery. The binding of these coregulatory proteins to the steroid hormone receptors is often mediated through their two activation functional domains, AF1, which resides in the N-terminal domain, and the ligand-dependent AF2, which is localized in the C-terminal ligand-binding domain. Compared to other important functional domains of the steroid hormone receptors, our understanding of the mechanisms of action of the AF1 are incomplete, in part, due to the fact that, in solution, AF1 is intrinsically disordered (ID). However, recent studies have shown that AF1 must adopt a functionally active and folded conformation for its optimal activity under physiological conditions. In this review, we summarize and discuss current knowledge regarding the molecular mechanisms of AF1-mediated gene activation, focusing on AF1 conformation and coactivator binding. We further propose models for the binding/folding of the AF1 domains of the steroid hormone receptors and their protein:protein interactions. The population of ID AF1 can be visualized as a collection of many different conformations, some of which may be assuming the proper functional folding for other critical target binding partners that result in the ultimate assembly of AF1:coactivator complexes and subsequent gene regulation. Knowledge of the mechanisms involved therein will significantly help in understanding how signals from a steroid to a specific target gene are conveyed.
Figures
Similar articles
-
Binding of the N-terminal region of coactivator TIF2 to the intrinsically disordered AF1 domain of the glucocorticoid receptor is accompanied by conformational reorganizations.J Biol Chem. 2012 Dec 28;287(53):44546-60. doi: 10.1074/jbc.M112.411330. Epub 2012 Nov 6. J Biol Chem. 2012. PMID: 23132854 Free PMC article.
-
TBP binding-induced folding of the glucocorticoid receptor AF1 domain facilitates its interaction with steroid receptor coactivator-1.PLoS One. 2011;6(7):e21939. doi: 10.1371/journal.pone.0021939. Epub 2011 Jul 7. PLoS One. 2011. PMID: 21760925 Free PMC article.
-
Binding-folding induced regulation of AF1 transactivation domain of the glucocorticoid receptor by a cofactor that binds to its DNA binding domain.PLoS One. 2011;6(10):e25875. doi: 10.1371/journal.pone.0025875. Epub 2011 Oct 7. PLoS One. 2011. PMID: 22003412 Free PMC article.
-
Variable steroid receptor responses: Intrinsically disordered AF1 is the key.Mol Cell Endocrinol. 2013 Aug 25;376(1-2):81-4. doi: 10.1016/j.mce.2013.06.007. Epub 2013 Jun 17. Mol Cell Endocrinol. 2013. PMID: 23792173 Free PMC article. Review.
-
Structure and function of steroid receptor AF1 transactivation domains: induction of active conformations.Biochem J. 2005 Nov 1;391(Pt 3):449-64. doi: 10.1042/BJ20050872. Biochem J. 2005. PMID: 16238547 Free PMC article. Review.
Cited by
-
Molecular mechanism of estrogen-estrogen receptor signaling.Reprod Med Biol. 2016 Dec 5;16(1):4-20. doi: 10.1002/rmb2.12006. eCollection 2017 Jan. Reprod Med Biol. 2016. PMID: 29259445 Free PMC article. Review.
-
Allosteric modulators of steroid hormone receptors: structural dynamics and gene regulation.Endocr Rev. 2012 Apr;33(2):271-99. doi: 10.1210/er.2011-1033. Epub 2012 Mar 20. Endocr Rev. 2012. PMID: 22433123 Free PMC article. Review.
-
Role of Steroid Hormones in the Pathogenesis of Nonalcoholic Fatty Liver Disease.Metabolites. 2021 May 17;11(5):320. doi: 10.3390/metabo11050320. Metabolites. 2021. PMID: 34067649 Free PMC article. Review.
-
Effect of the Use of Gnrh Analogs in Low-Grade Cerebral Glioma.Children (Basel). 2023 Jan 5;10(1):115. doi: 10.3390/children10010115. Children (Basel). 2023. PMID: 36670665 Free PMC article.
-
An intrinsically disordered region-mediated confinement state contributes to the dynamics and function of transcription factors.Mol Cell. 2021 Apr 1;81(7):1484-1498.e6. doi: 10.1016/j.molcel.2021.01.013. Epub 2021 Feb 8. Mol Cell. 2021. PMID: 33561389 Free PMC article.
References
-
- Beato M, Klug J. Steroid hormone receptors: an update. Human reproduction update. 2000;6:225–236. - PubMed
-
- Chawla A, Repa JJ, Evans RM, Mangelsdorf DJ. Nuclear receptors and lipid physiology: opening the X-files. Science. 2001;294:1866–1870. - PubMed
-
- Li X, O'Malley BW. Unfolding the action of progesterone receptors. J Biol Chem. 2003;278:39261–39264. - PubMed
-
- Lonard DM, O'Malley BW. Nuclear receptor coregulators: judges, juries, and executioners of cellular regulation. Mol Cell. 2007;27:691–700. - PubMed
-
- Kumar R, Thompson EB. The structure of the nuclear hormone receptors. Steroids. 1999;64:310–319. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
