

Critical role for tetrahydrobiopterin recycling by dihydrofolate reductase in regulation of endothelial nitric-oxide synthase coupling: relative importance of the de novo biopterin synthesis versus salvage pathways
- PMID: 19666465
- PMCID: PMC2788863
- DOI: 10.1074/jbc.M109.041483
Critical role for tetrahydrobiopterin recycling by dihydrofolate reductase in regulation of endothelial nitric-oxide synthase coupling: relative importance of the de novo biopterin synthesis versus salvage pathways
Abstract
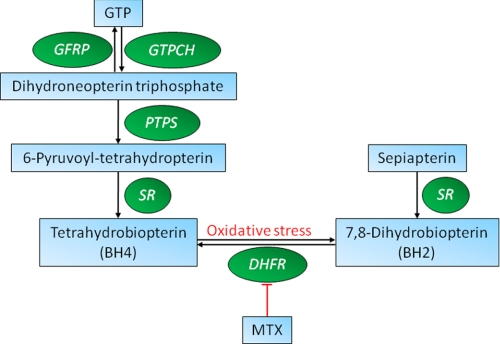
Tetrahyrobiopterin (BH4) is a required cofactor for the synthesis of nitric oxide by endothelial nitric-oxide synthase (eNOS), and BH4 bioavailability within the endothelium is a critical factor in regulating the balance between NO and superoxide production by eNOS (eNOS coupling). BH4 levels are determined by the activity of GTP cyclohydrolase I (GTPCH), the rate-limiting enzyme in de novo BH4 biosynthesis. However, BH4 levels may also be influenced by oxidation, forming 7,8-dihydrobiopterin (BH2), which promotes eNOS uncoupling. Conversely, dihydrofolate reductase (DHFR) can regenerate BH4 from BH2, but the functional importance of DHFR in maintaining eNOS coupling remains unclear. We investigated the role of DHFR in regulating BH4 versus BH2 levels in endothelial cells and in cell lines expressing eNOS combined with tet-regulated GTPCH expression in order to compare the effects of low or high levels of de novo BH4 biosynthesis. Pharmacological inhibition of DHFR activity by methotrexate or genetic knockdown of DHFR protein by RNA interference reduced intracellular BH4 and increased BH2 levels resulting in enzymatic uncoupling of eNOS, as indicated by increased eNOS-dependent superoxide but reduced NO production. In contrast to the decreased BH4:BH2 ratio induced by DHFR knockdown, GTPCH knockdown greatly reduced total biopterin levels but with no change in BH4:BH2 ratio. In cells expressing eNOS with low biopterin levels, DHFR inhibition or knockdown further diminished the BH4:BH2 ratio and exacerbated eNOS uncoupling. Taken together, these data reveal a key role for DHFR in eNOS coupling by maintaining the BH4:BH2 ratio, particularly in conditions of low total biopterin availability.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
