

TRPA channels distinguish gravity sensing from hearing in Johnston's organ
- PMID: 19666538
- PMCID: PMC2717111
- DOI: 10.1073/pnas.0906377106
TRPA channels distinguish gravity sensing from hearing in Johnston's organ
Abstract
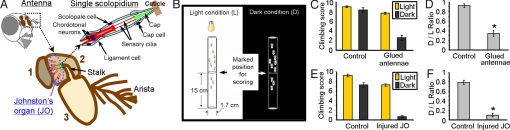
Although many animal species sense gravity for spatial orientation, the molecular bases remain uncertain. Therefore, we studied Drosophila melanogaster, which possess an inherent upward movement against gravity-negative geotaxis. Negative geotaxis requires Johnston's organ, a mechanosensory structure located in the antenna that also detects near-field sound. Because channels of the transient receptor potential (TRP) superfamily can contribute to mechanosensory signaling, we asked whether they are important for negative geotaxis. We identified distinct expression patterns for 5 TRP genes; the TRPV genes nanchung and inactive were present in most Johnston's organ neurons, the TRPN gene nompC and the TRPA gene painless were localized to 2 subpopulations of neurons, and the TRPA gene pyrexia was expressed in cap cells that may interact with the neurons. Likewise, mutating specific TRP genes produced distinct phenotypes, disrupting negative geotaxis (painless and pyrexia), hearing (nompC), or both (nanchung and inactive). Our genetic, physiological and behavioral data indicate that the sensory component of negative geotaxis involves multiple TRP genes. The results also distinguish between different mechanosensory modalities and set the stage for understanding how TRP channels contribute to mechanosensation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Kamikouchi A, et al. The neural basis of Drosophila gravity-sensing and hearing. Nature. 2009;458:165–171. - PubMed
-
- Kamikouchi A, Shimada T, Ito K. Comprehensive classification of the auditory sensory projections in the brain of the fruit fly Drosophila melanogaster. J Comp Neurol. 2006;499:317–356. - PubMed
-
- Yack JE. The structure and function of auditory chordotonal organs in insects. Microsc Res Tech. 2004;63:315–337. - PubMed
-
- Gopfert MC, Robert D. The mechanical basis of Drosophila audition. J Exp Biol. 2002;205:1199–1208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
