

Darwinian selection for sites of Asn-linked glycosylation in phylogenetically disparate eukaryotes and viruses
- PMID: 19666543
- PMCID: PMC2726397
- DOI: 10.1073/pnas.0905818106
Darwinian selection for sites of Asn-linked glycosylation in phylogenetically disparate eukaryotes and viruses
Abstract
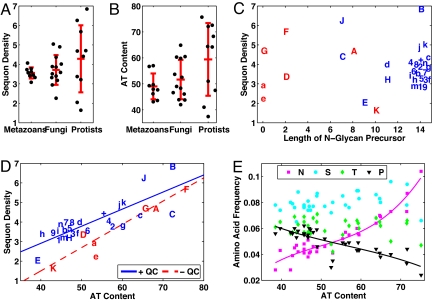
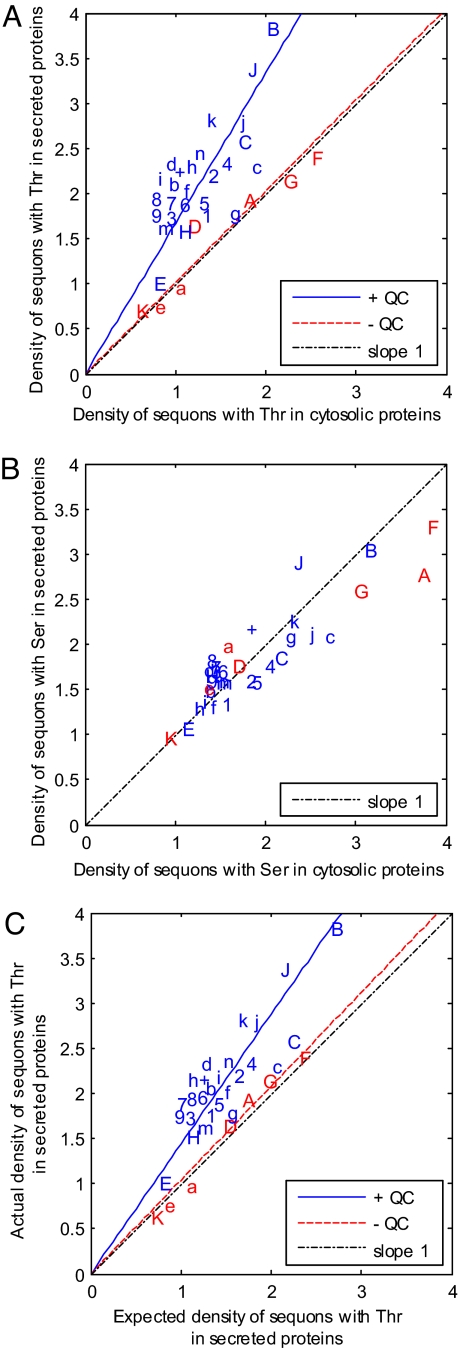
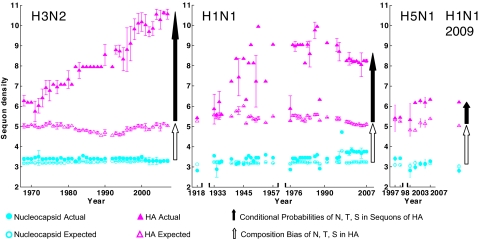
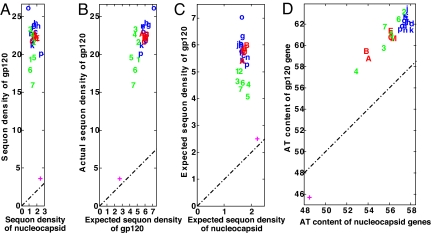
Numerous protists and rare fungi have truncated Asn-linked glycan precursors and lack N-glycan-dependent quality control (QC) systems for glycoprotein folding in the endoplasmic reticulum. Here, we show that the abundance of sequons (NXT or NXS), which are sites for N-glycosylation of secreted and membrane proteins, varies by more than a factor of 4 among phylogenetically diverse eukaryotes, based on a few variables. There is positive correlation between the density of sequons and the AT content of coding regions, although no causality can be inferred. In contrast, there appears to be Darwinian selection for sequons containing Thr, but not Ser, in eukaryotes that have N-glycan-dependent QC systems. Selection for sequons with Thr, which nearly doubles the sequon density in human secreted and membrane proteins, occurs by an increased conditional probability that Asn and Thr are present in sequons rather than elsewhere. Increasing sequon densities of the hemagglutinin (HA) of influenza viruses A/H3N2 and A/H1N1 during the past few decades of human infection also result from an increased conditional probability that Asn, Thr, and Ser are present in sequons rather than elsewhere. In contrast, there is no selection on sequons by this mechanism in HA of A/H5N1 or 2009 A/H1N1 (Swine flu). Very strong selection for sequons with both Thr and Ser in glycoprotein of M(r) 120,000 (gp120) of HIV and related retroviruses results from this same mechanism, as well as amino acid composition bias and increases in AT content. We conclude that there is Darwinian selection for sequons in phylogenetically disparate eukaryotes and viruses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Helenius A, Aebi M. Roles of N-linked glycans in the endoplasmic reticulum. Annu Rev Biochem. 2004;73:1019–1049. - PubMed
-
- Kornfeld R, Kornfeld S. Assembly of asparagine-linked oligosaccharides. Annu Rev Biochem. 1985;54:631–664. - PubMed
-
- Apweiler R, Hermjakob H, Sharon N. On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim Biophys Acta. 1999;1473:4–8. - PubMed
-
- Ben-Dor S, Esterman N, Rubin E, Sharon N. Biases and complex patterns in the residues flanking protein N-glycosylation sites. Glycobiology. 2004;14:95–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
