

Phospholipid meets all-trans-retinal: the making of RPE bisretinoids
- PMID: 19666736
- PMCID: PMC2803227
- DOI: 10.1194/jlr.R000687
Phospholipid meets all-trans-retinal: the making of RPE bisretinoids
Abstract
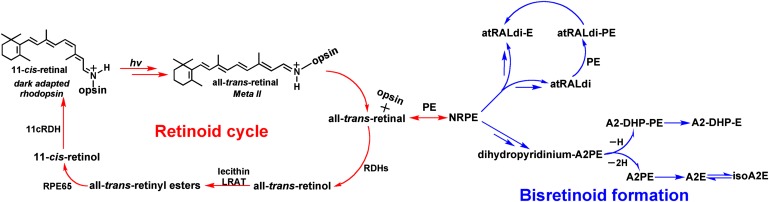
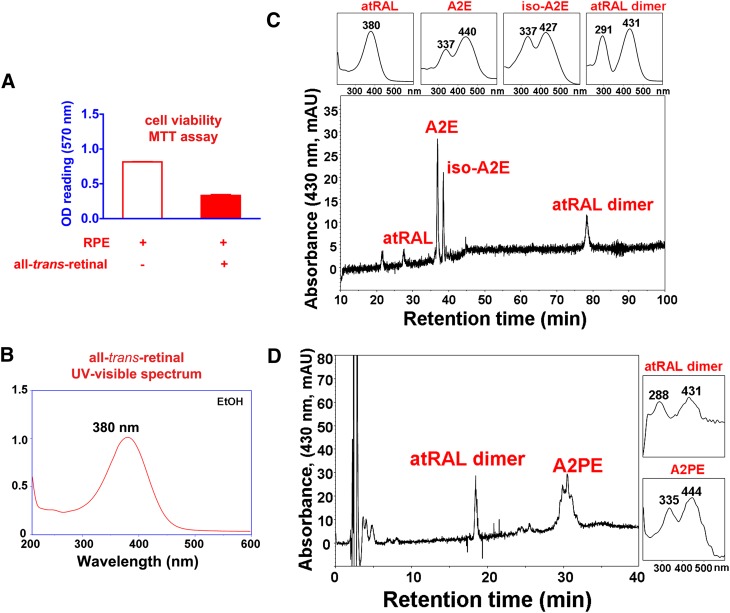
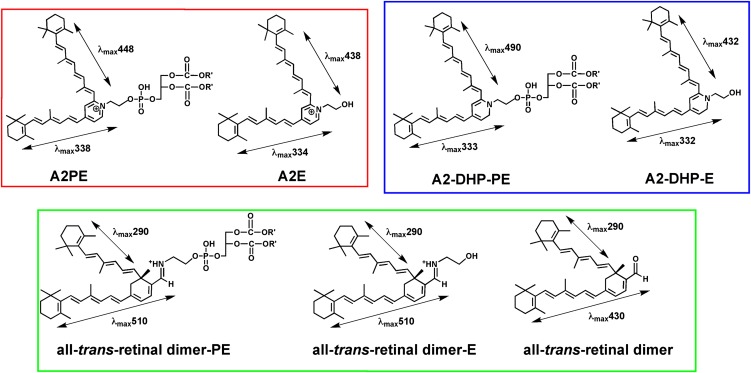
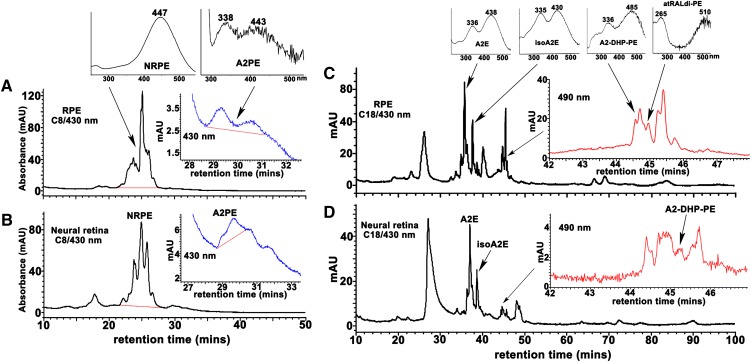
The lipid phase of the photoreceptor outer segment membrane is essential to the photon capturing and signaling functions of rhodopsin. Rearrangement of phospholipids in the bilayer accompanies the formation of the active intermediates of rhodopsin following photon absorption. Furthermore, evidence for the formation of a condensation product between the photolyzed chromophore all-trans-retinal and phosphatidylethanolamine indicates that phospholipid may also participate in the movement of the retinoid in the membrane. The downside of these interactions is the formation of bisretinoid-phosphatidylethanolamine compounds that accumulate in retinal pigment epithelial cells with age and that are particularly abundant in some retinal disorders. The propensity of these compounds to negatively impact on the cells has been linked to the pathogenesis of some retinal disorders including juvenile onset recessive Stargardt disease and age-related macular degeneration.
Figures
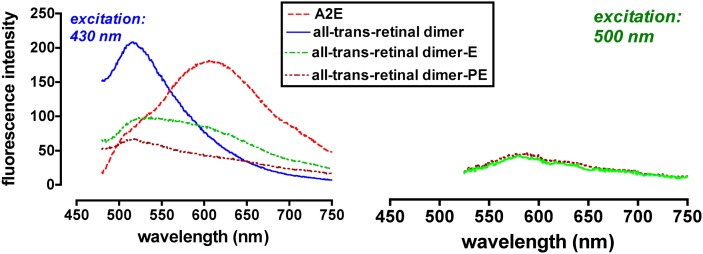
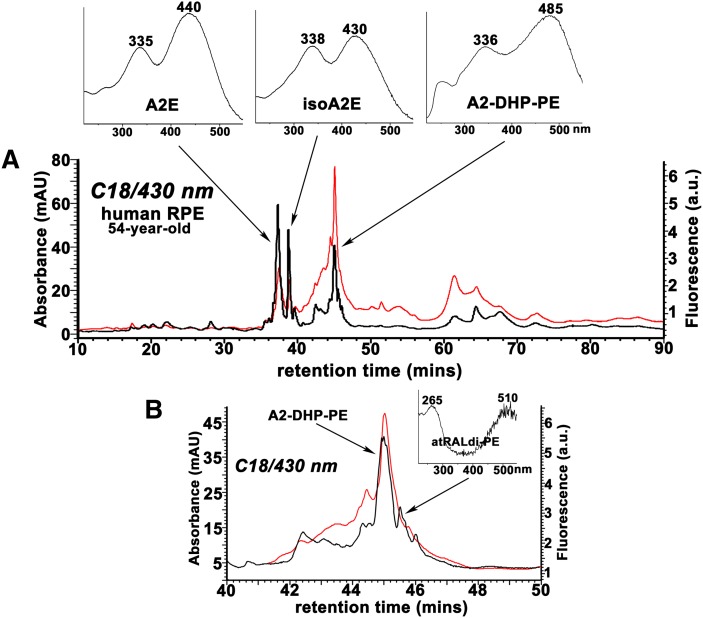
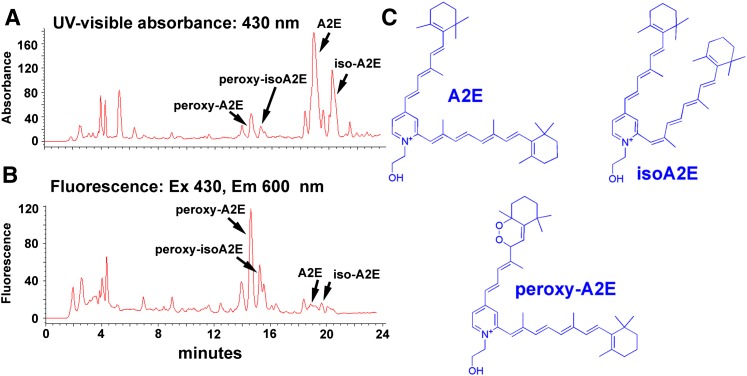
References
-
- Hicks D., Sparrow J. R., Barnstable C. J. 1989. Immunoelectron microscopical examination of the surface distribution of opsin in rat rod photoreceptor cells. Exp. Eye Res. 49: 13–29. - PubMed
-
- Fliesler S. J., Anderson R. E. 1983. Chemistry and metabolism of lipids in the vertebrate retina. Prog. Lipid Res. 22: 79–131. - PubMed
-
- McBee J. K., Palczewski K., Baehr W., Pepperberg D. R. 2001. Confronting complexity: the interlink of phototransduction and retinoid metabolism in the vertebrate retina. Prog. Retin. Eye Res. 20: 469–529. - PubMed
-
- Beck M., Siebert F., Sakmar T. P. 1998. Evidence for the specific interaction of a lipid molecule with rhodopsin which is altered in the transition to the active state metarhodopsin II. FEBS Lett. 436: 304–308. - PubMed
-
- Hessel E., Muller P., Hermann A., Hofmann K. P. 2001. Light-induced reorganization of phospholipid in rod disc membrane. J. Biol. Chem. 276: 2538–2543. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
