

Abscisic acid has contrasting effects on salt excretion and polyamine concentrations of an inland and a coastal population of the Mediterranean xero-halophyte species Atriplex halimus
- PMID: 19666900
- PMCID: PMC2749539
- DOI: 10.1093/aob/mcp174
Abscisic acid has contrasting effects on salt excretion and polyamine concentrations of an inland and a coastal population of the Mediterranean xero-halophyte species Atriplex halimus
Abstract
Background and aims: Different populations of the Mediterranean xerohalophyte species Atriplex halimus exhibit different levels of resistance to salt and osmotic stress depending on the nature of the osmocompatible solute they accumulate. There is, however, no conclusive description of the involvement of abscisic acid (ABA) in the plant response to NaCl or osmotic stress in this species.
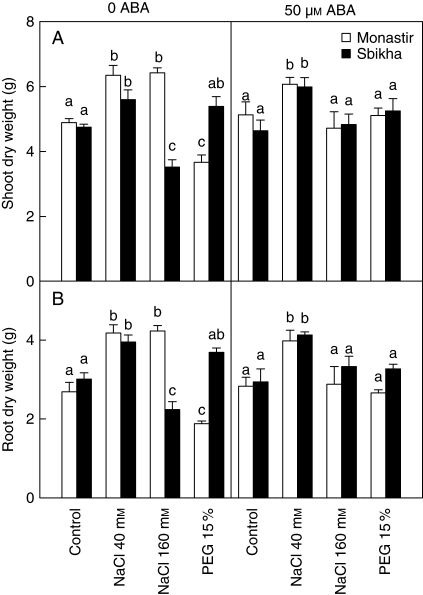
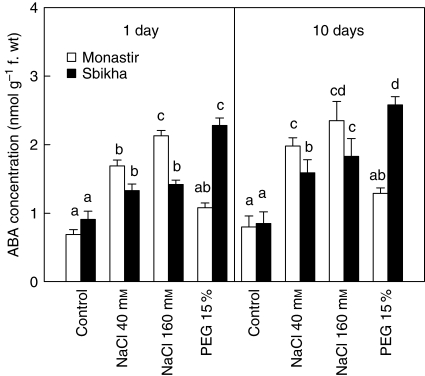
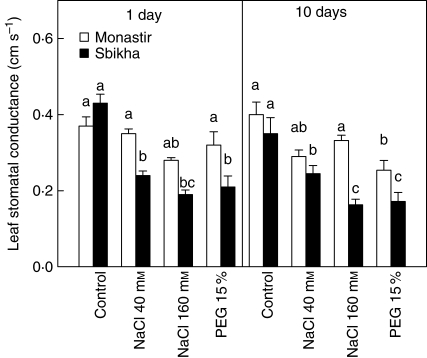
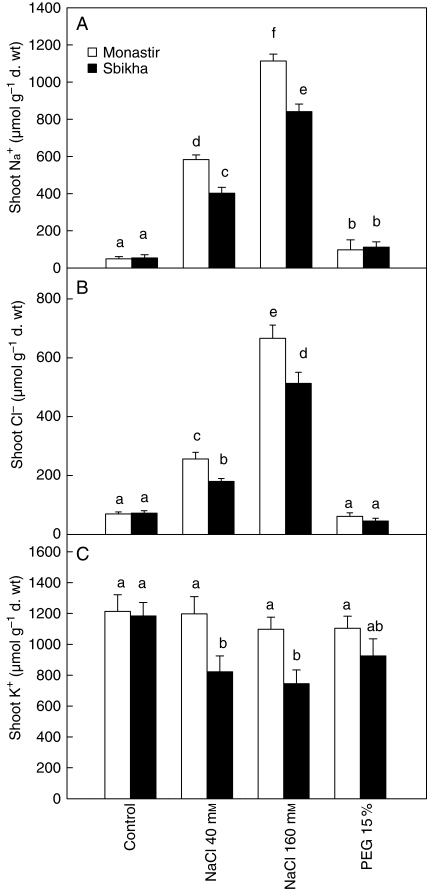
Methods: Seedlings issued from an inland water-stress-resistant population (Sbikha) and from a coastal salt-resistant one (Monastir) were exposed in nutrient solutions to NaCl (40 or 160 mm) or to 15 % PEG for 1 d and 10 d in the presence or absence of 50 microm ABA.
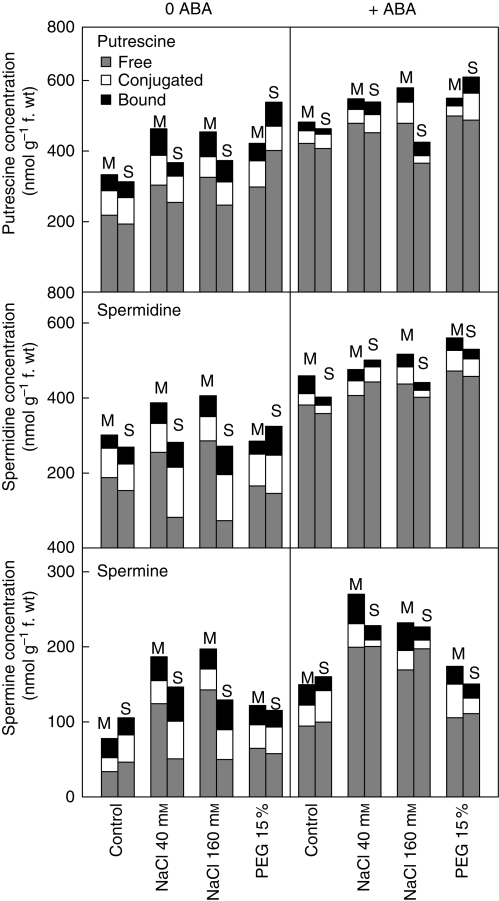
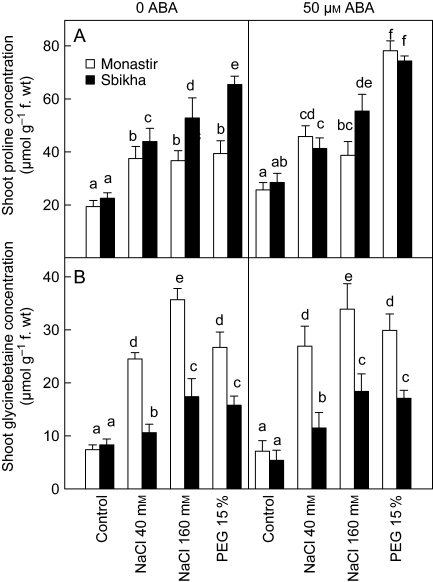
Key results: Plants from Sbikha accumulated higher amounts of ABA in response to osmotic stress than those of Monastir, while an opposite trend was recorded for NaCl exposure. Exogenous ABA improved osmotic stress resistance in Monastir through an improvement in the efficiency of stomatal conductance regulation. It also improved NaCl resistance in Sbikha through an increase in sodium excretion through the external bladders. It is suggested that polyamines (spermidine and spermine) are involved in the salt excretion process and that ABA contributes to polyamine synthesis as well as to the conversion from the bound and conjugated to the free soluble forms of polyamine. Proline accumulated in response to osmotic stress and slightly increased in response to ABA treatment while glycinebetaine accumulated in response to salinity and was not influenced by ABA.
Conclusions: It is concluded that ABA is involved in both salt and osmotic stress resistance in the xerohalophyte species Atriplex halimus but that it acts on different physiological cues in response to those distinct environmental constraints.
Figures

References
-
- Alcázar R, Cuevas JC, Patron M, ltabella T, Tiburcio AF. Abscisic acid modulates polyamine metabolism under water stress in Arabidopsis thaliana. Physiologia Plantarum. 2006;128:448–455.
-
- Antolín MC, Santesteban HY, Santa María E, Aguirreolea J, Sánchez-Díaz M. Involvement of abscisic acid and polyamines in berry ripening of Vitis vinifera (L.) subjected to water deficit irrigation. Australian Journal of Grape and Wine Research. 2008;114:123–133.
-
- Arena ME, Pastur GM, Benavides MP, Curvetto N. Polyamines and inhibitors used in successive culture media for in vitro rooting in Berberis buxifolia. New Zealand Journal of Botany. 2005;43:373–380.
-
- Aslam Z, Jeschke WD, Barrettlennard EG, Setter TL, Watkin E, Greenway H. Effect of external NaCl on the growth of Atriplex amnicola and the ion relations and carbohydrate status of the leaves. Plant, Cell & Environment. 1986;9:571–580.
-
- Aurisano N, Bertani A, Mattana M, Reggiani R. Abscisic-acid induced stress-like polyamine pattern in wheat seedlings and its reversal by potassium ions. Physiologia Plantarum. 1993;89:687–692.
