

Cell cycle-dependent role of MRN at dysfunctional telomeres: ATM signaling-dependent induction of nonhomologous end joining (NHEJ) in G1 and resection-mediated inhibition of NHEJ in G2
- PMID: 19667071
- PMCID: PMC2756883
- DOI: 10.1128/MCB.00476-09
Cell cycle-dependent role of MRN at dysfunctional telomeres: ATM signaling-dependent induction of nonhomologous end joining (NHEJ) in G1 and resection-mediated inhibition of NHEJ in G2
Abstract
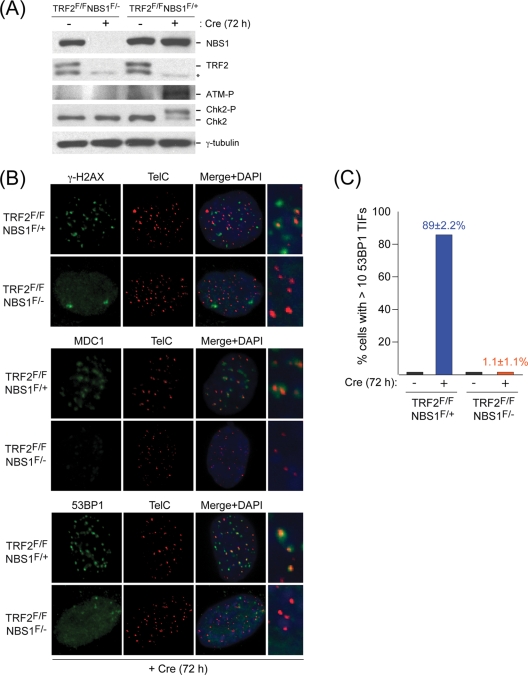
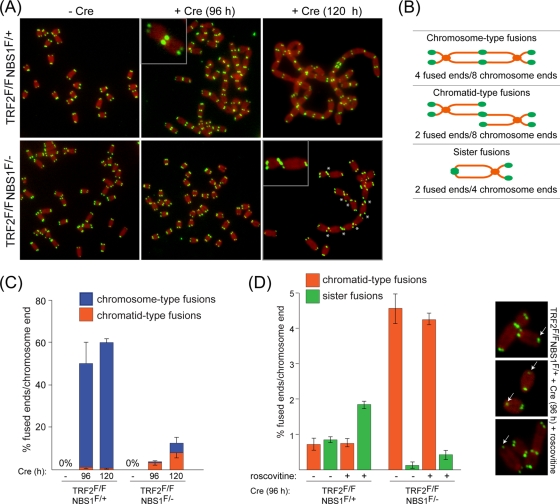
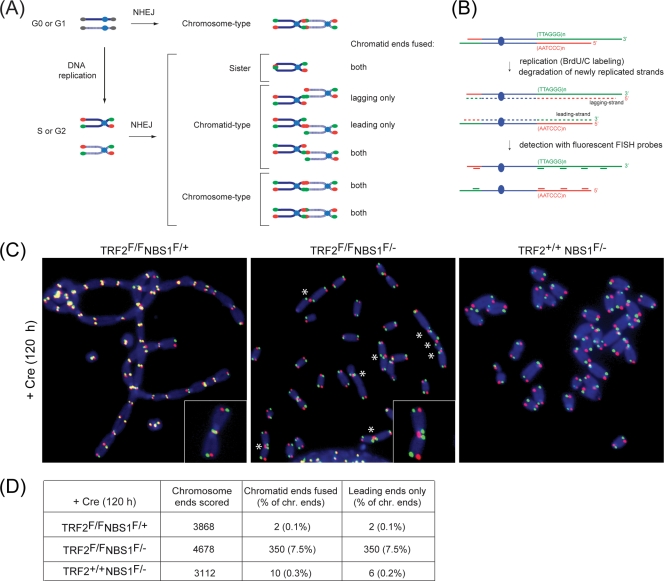
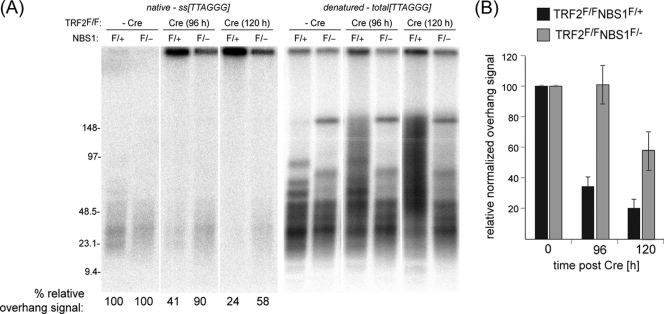
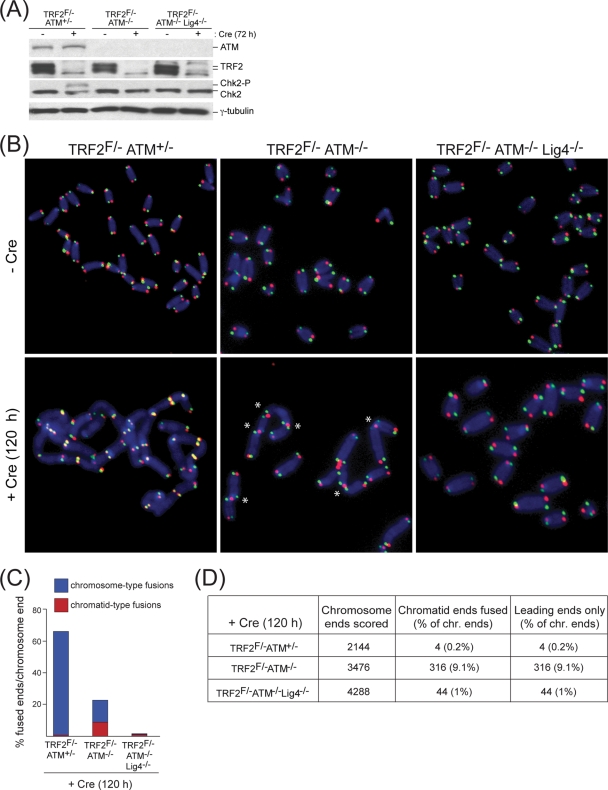
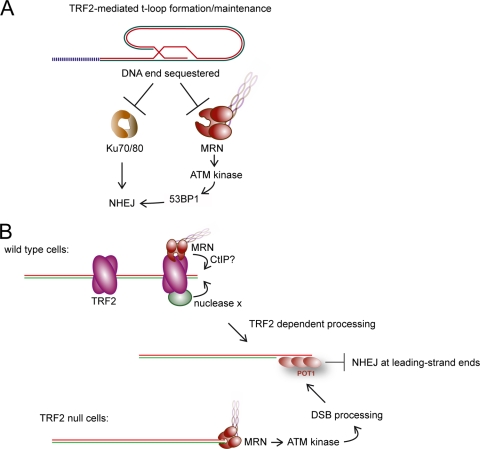
Here, we address the role of the MRN (Mre11/Rad50/Nbs1) complex in the response to telomeres rendered dysfunctional by deletion of the shelterin component TRF2. Using conditional NBS1/TRF2 double-knockout MEFs, we show that MRN is required for ATM signaling in response to telomere dysfunction. This establishes that MRN is the only sensor for the ATM kinase and suggests that TRF2 might block ATM signaling by interfering with MRN binding to the telomere terminus, possibly by sequestering the telomere end in the t-loop structure. We also examined the role of the MRN/ATM pathway in nonhomologous end joining (NHEJ) of damaged telomeres. NBS1 deficiency abrogated the telomere fusions that occur in G(1), consistent with the requirement for ATM and its target 53BP1 in this setting. Interestingly, NBS1 and ATM, but not H2AX, repressed NHEJ at dysfunctional telomeres in G(2), specifically at telomeres generated by leading-strand DNA synthesis. Leading-strand telomere ends were not prone to fuse in the absence of either TRF2 or MRN/ATM, indicating redundancy in their protection. We propose that MRN represses NHEJ by promoting the generation of a 3' overhang after completion of leading-strand DNA synthesis. TRF2 may ensure overhang formation by recruiting MRN (and other nucleases) to newly generated telomere ends. The activation of the MRN/ATM pathway by the dysfunctional telomeres is proposed to induce resection that protects the leading-strand ends from NHEJ when TRF2 is absent. Thus, the role of MRN at dysfunctional telomeres is multifaceted, involving both repression of NHEJ in G(2) through end resection and induction of NHEJ in G(1) through ATM-dependent signaling.
Figures
References
-
- Bai, Y., and J. P. Murnane. 2003. Telomere instability in a human tumor cell line expressing NBS1 with mutations at sites phosphorylated by ATM. Mol. Cancer Res. 1:1058-1069. - PubMed
-
- Bailey, S. M., M. N. Cornforth, A. Kurimasa, D. J. Chen, and E. H. Goodwin. 2001. Strand-specific postreplicative processing of mammalian telomeres. Science 293:2462-2465. - PubMed
-
- Bailey, S. M., E. H. Goodwin, and M. N. Cornforth. 2004. Strand-specific fluorescence in situ hybridization: the CO-FISH family. Cytogenet. Genome Res. 107:14-17. - PubMed
-
- Bailey, S. M., M. A. Brenneman, J. Halbrook, J. A. Nickoloff, R. L. Ullrich, and E. H. Goodwin. 2004. The kinase activity of DNA-PK is required to protect mammalian telomeres. DNA Repair (Amsterdam) 3:225-233. - PubMed
-
- Bassing, C. H., K. F. Chua, J. Sekiguchi, H. Suh, S. R. Whitlow, J. C. Fleming, B. C. Monroe, D. N. Ciccone, C. Yan, K. Vlasakova, D. M. Livingston, D. O. Ferguson, R. Scully, and F. W. Alt. 2002. Increased ionizing radiation sensitivity and genomic instability in the absence of histone H2AX. Proc. Natl. Acad. Sci. USA 99:8173-8178. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous