

FOXC1 is required for normal cerebellar development and is a major contributor to chromosome 6p25.3 Dandy-Walker malformation
- PMID: 19668217
- PMCID: PMC2843139
- DOI: 10.1038/ng.422
FOXC1 is required for normal cerebellar development and is a major contributor to chromosome 6p25.3 Dandy-Walker malformation
Abstract
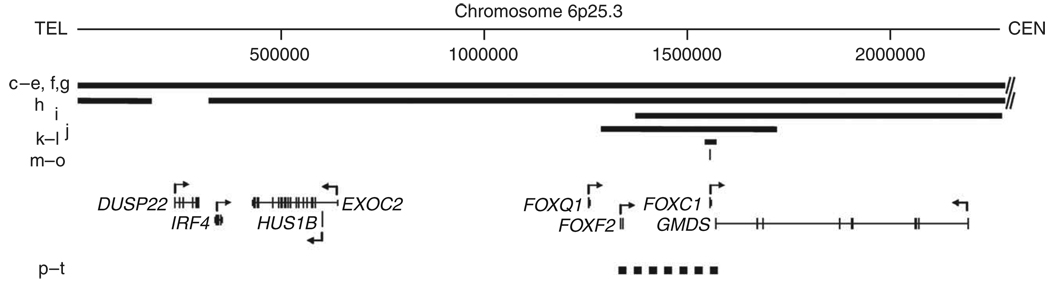
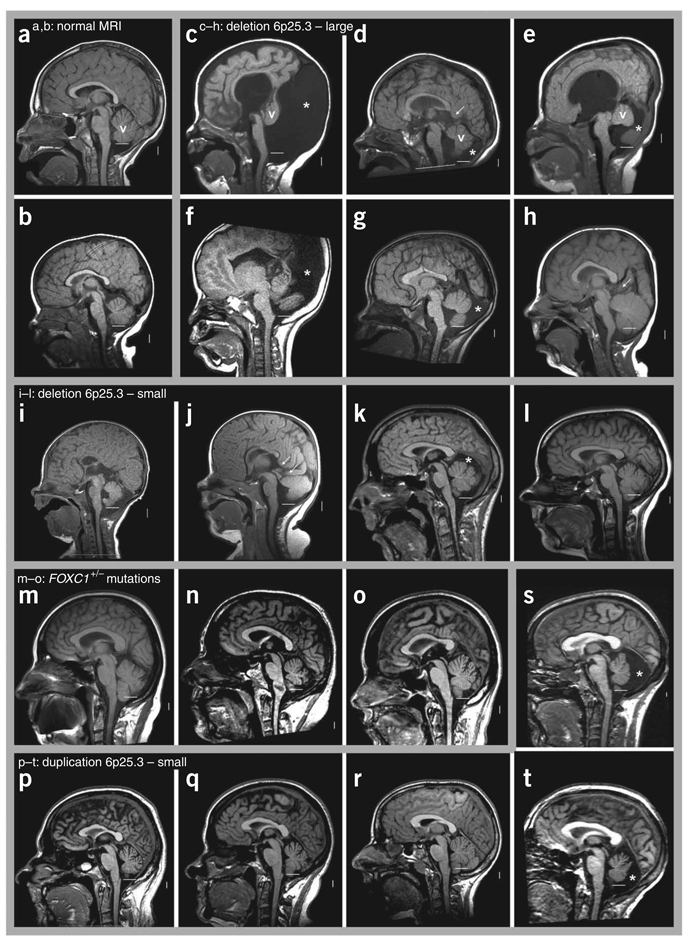
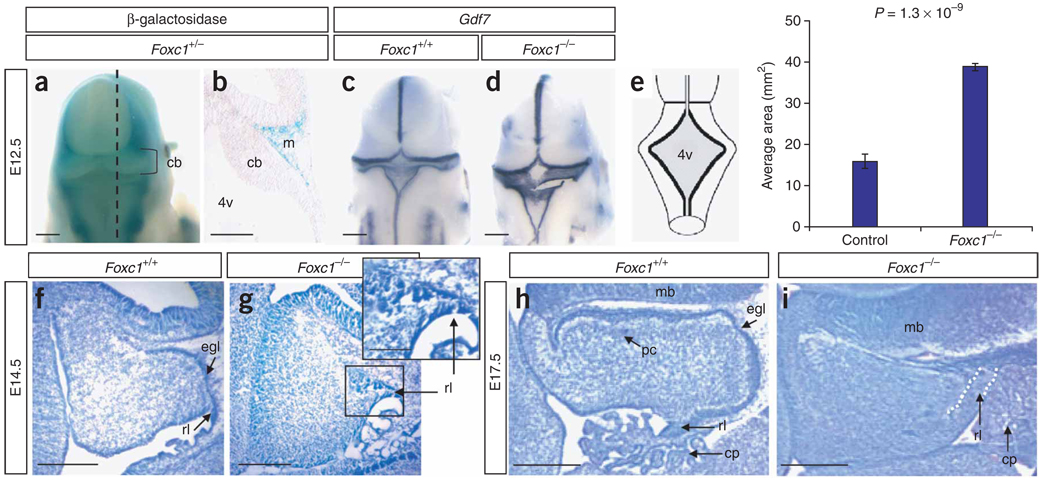
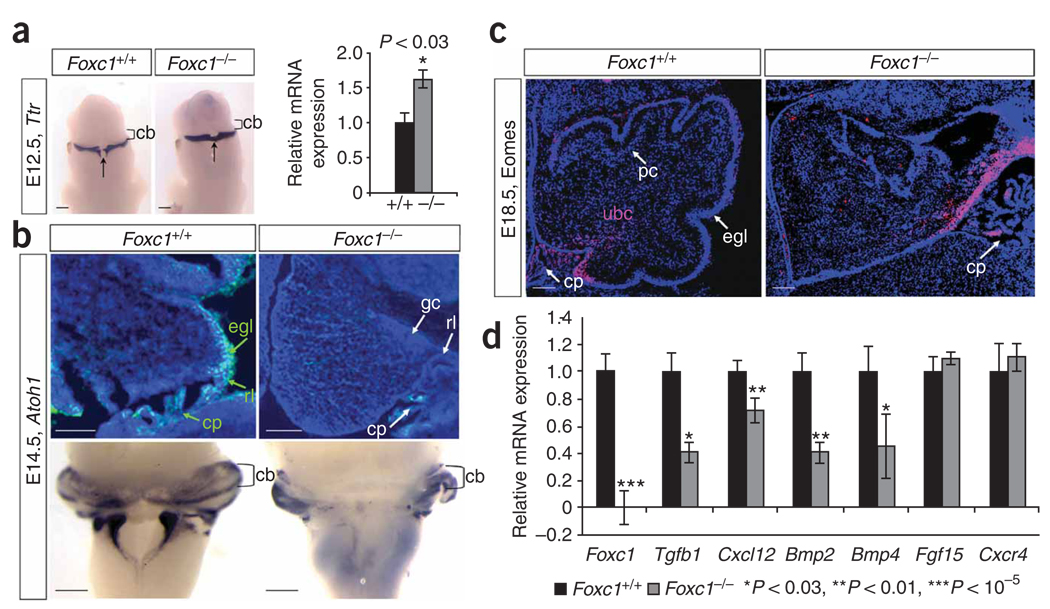
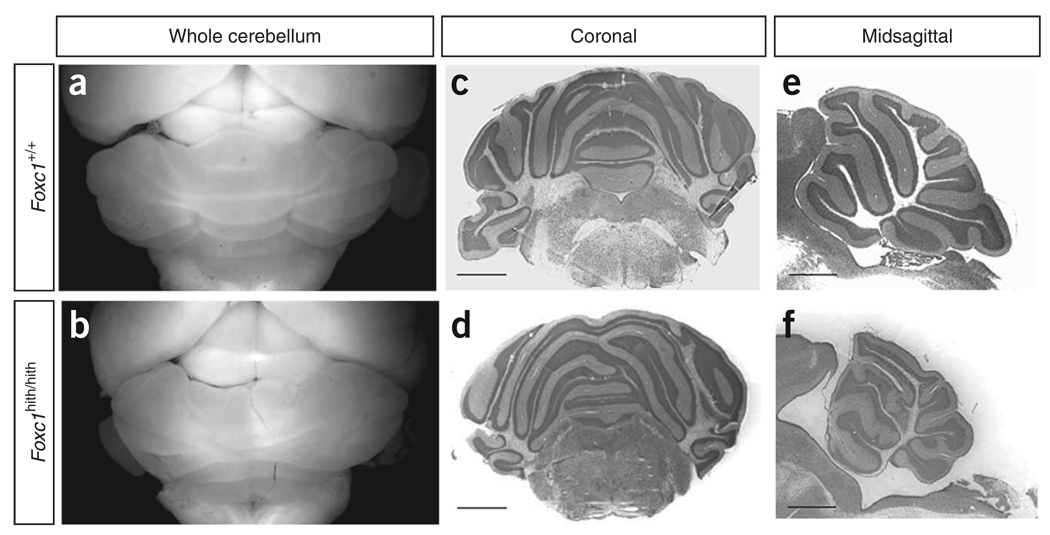
Dandy-Walker malformation (DWM), the most common human cerebellar malformation, has only one characterized associated locus. Here we characterize a second DWM-linked locus on 6p25.3, showing that deletions or duplications encompassing FOXC1 are associated with cerebellar and posterior fossa malformations including cerebellar vermis hypoplasia (CVH), mega-cisterna magna (MCM) and DWM. Foxc1-null mice have embryonic abnormalities of the rhombic lip due to loss of mesenchyme-secreted signaling molecules with subsequent loss of Atoh1 expression in vermis. Foxc1 homozygous hypomorphs have CVH with medial fusion and foliation defects. Human FOXC1 heterozygous mutations are known to affect eye development, causing a spectrum of glaucoma-associated anomalies (Axenfeld-Rieger syndrome, ARS; MIM no. 601631). We report the first brain imaging data from humans with FOXC1 mutations and show that these individuals also have CVH. We conclude that alteration of FOXC1 function alone causes CVH and contributes to MCM and DWM. Our results highlight a previously unrecognized role for mesenchyme-neuroepithelium interactions in the mid-hindbrain during early embryogenesis.
Figures
References
-
- Grinberg I, et al. Heterozygous deletion of the linked genes ZIC1 and ZIC4 is involved in Dandy-Walker malformation. Nat. Genet. 2004;36:1053–1055. - PubMed
-
- Parisi MA, Dobyns WB. Human malformations of the midbrain and hindbrain: review and proposed classification scheme. Mol. Genet. Metab. 2003;80:36–53. - PubMed
-
- DeScipio C, et al. Subtelomeric deletions of chromosome 6p: molecular and cytogenetic characterization of three new cases with phenotypic overlap with RitscherSchinzel (3C) syndrome. Am. J. Med. Genet. A. 2005;134:3–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
