

TANK is a negative regulator of Toll-like receptor signaling and is critical for the prevention of autoimmune nephritis
- PMID: 19668221
- PMCID: PMC2910115
- DOI: 10.1038/ni.1771
TANK is a negative regulator of Toll-like receptor signaling and is critical for the prevention of autoimmune nephritis
Abstract
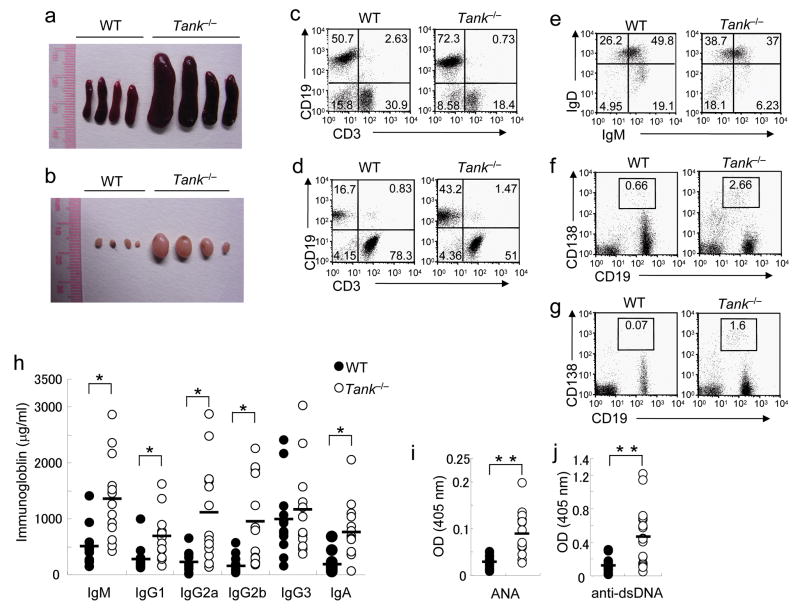
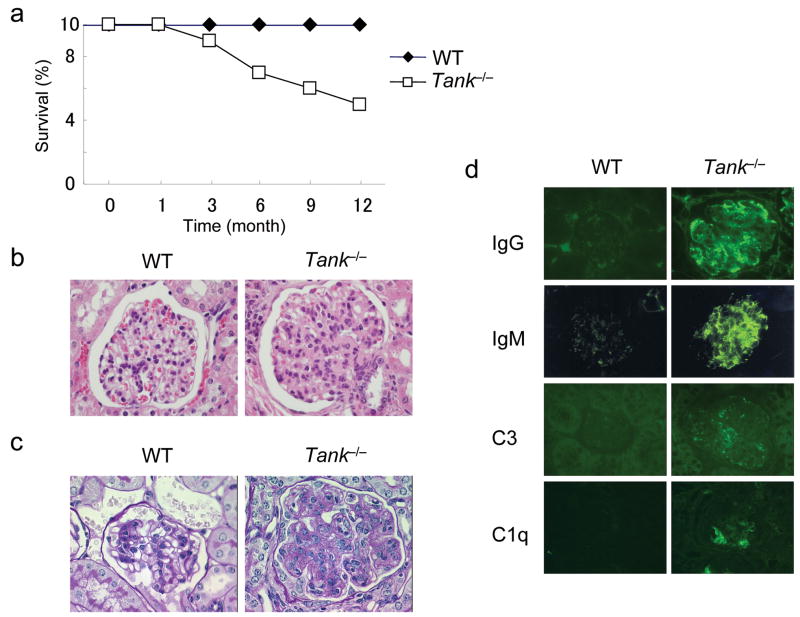
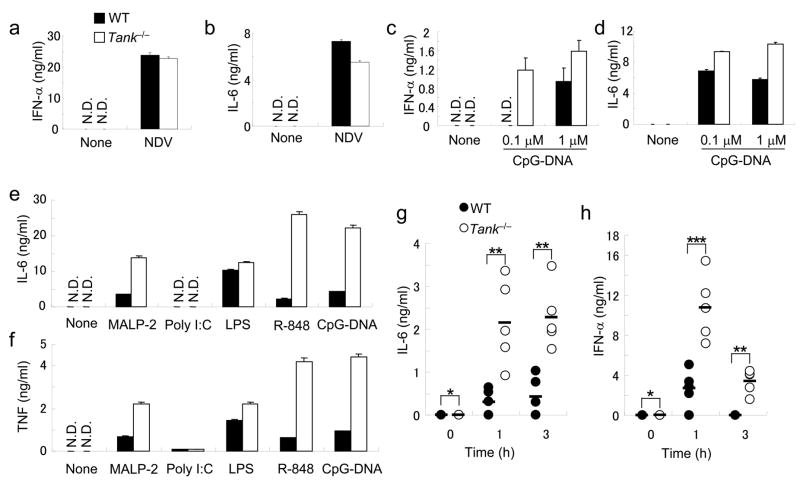
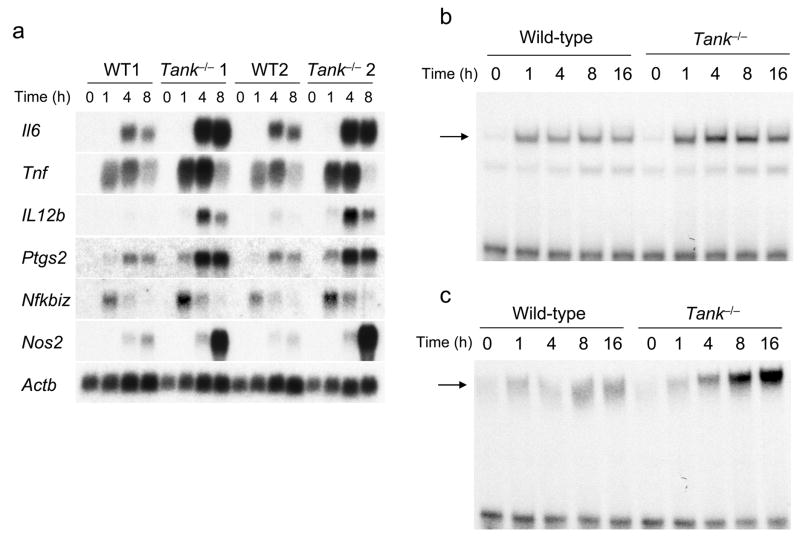
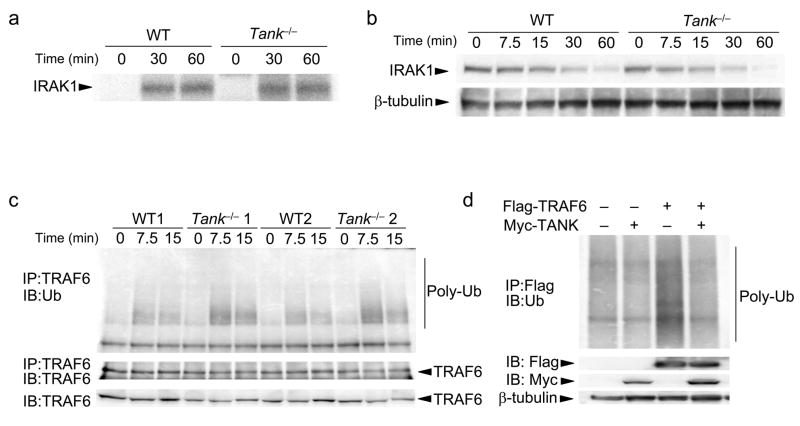
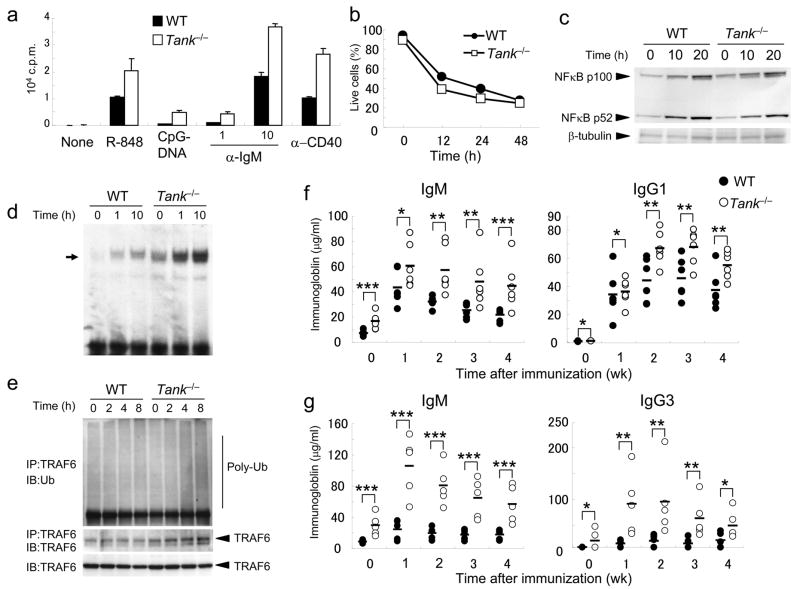
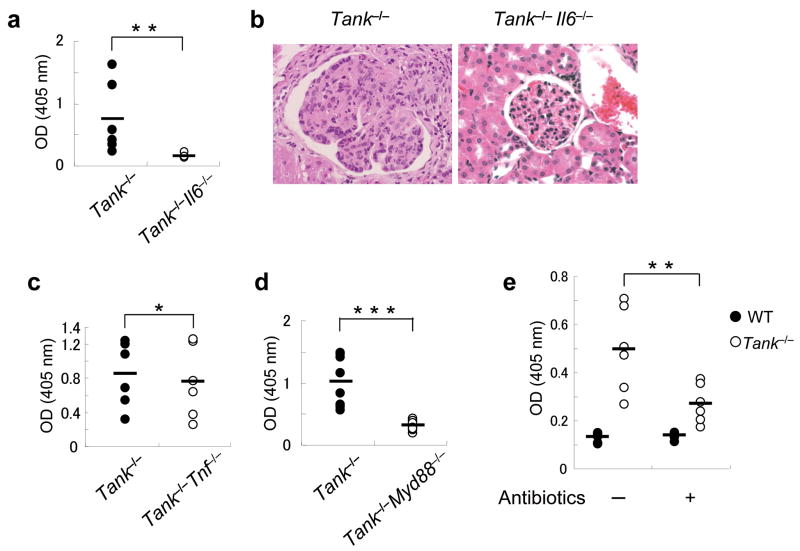
The intensity and duration of immune responses are controlled by many proteins that modulate Toll-like receptor (TLR) signaling. TANK has been linked to positive regulation of the transcription factors IRF3 and NF-kappaB. Here we demonstrate that TANK is not involved in interferon responses and is a negative regulator of proinflammatory cytokine production induced by TLR signaling. TLR-induced polyubiquitination of the ubiquitin ligase TRAF6 was upregulated in Tank(-/-) macrophages. Notably, Tank(-/-) mice spontaneously developed fatal glomerulonephritis owing to deposition of immune complexes. Autoantibody production in Tank(-/-) mice was abrogated by antibiotic treatment or the absence of interleukin 6 (IL-6) or the adaptor MyD88. Our results demonstrate that constitutive TLR signaling by intestinal commensal microflora is suppressed by TANK.
Conflict of interest statement
Disclosure
The authors declare that they have no competing financial interests.
Figures
References
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- Beutler B. Inferences, questions and possibilities in Toll-like receptor signalling. Nature. 2004;430:257–263. - PubMed
-
- Medzhitov R. Recognition of microorganisms and activation of the immune response. Nature. 2007;449:819–826. - PubMed
-
- Kawai T, Adachi O, Ogawa T, Takeda K, Akira S. Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity. 1999;11:115–122. - PubMed
-
- Yamamoto M, et al. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science. 2003;301:640–643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
