

Oscillations and synchrony in large-scale cortical network models
- PMID: 19669478
- PMCID: PMC2585628
- DOI: 10.1007/s10867-008-9079-y
Oscillations and synchrony in large-scale cortical network models
Abstract
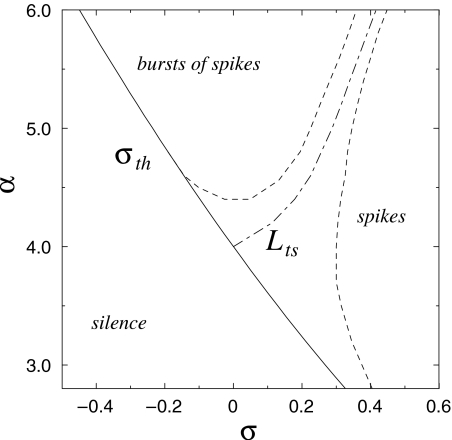
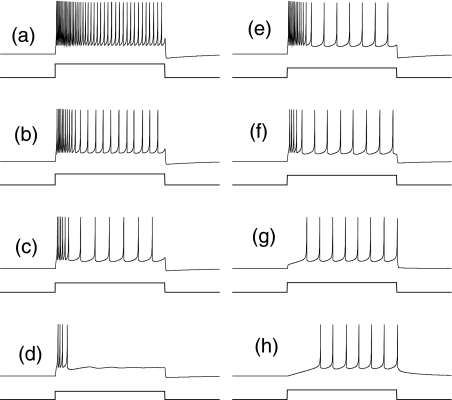
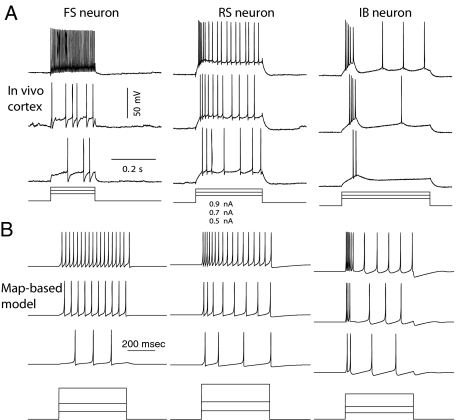
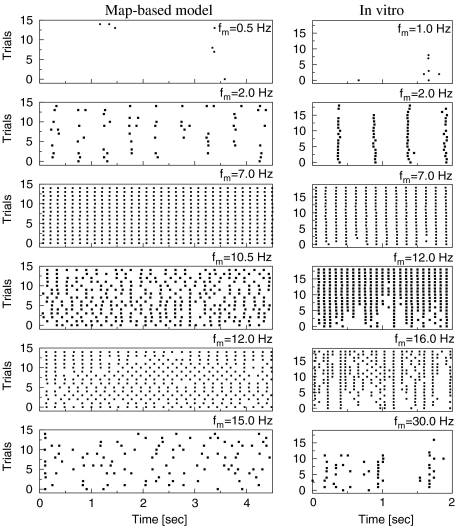
Intrinsic neuronal and circuit properties control the responses of large ensembles of neurons by creating spatiotemporal patterns of activity that are used for sensory processing, memory formation, and other cognitive tasks. The modeling of such systems requires computationally efficient single-neuron models capable of displaying realistic response properties. We developed a set of reduced models based on difference equations (map-based models) to simulate the intrinsic dynamics of biological neurons. These phenomenological models were designed to capture the main response properties of specific types of neurons while ensuring realistic model behavior across a sufficient dynamic range of inputs. This approach allows for fast simulations and efficient parameter space analysis of networks containing hundreds of thousands of neurons of different types using a conventional workstation. Drawing on results obtained using large-scale networks of map-based neurons, we discuss spatiotemporal cortical network dynamics as a function of parameters that affect synaptic interactions and intrinsic states of the neurons.
Figures











References
-
- Golomb, D.: Models of neuronal transient synchrony during propagation of activity through neocortical circuitry. J. Neurophysiol. 79, 1–12 (1998) - PubMed
-
- Golomb, D., Amitai, Y.: Propagating neuronal discharges in neocortical slices: computational and experimental study. J. Neurophysiol. 78, 1199–1211 (1997) - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
