

Alteration of oligomeric state and domain architecture is essential for functional transformation between transferase and hydrolase with the same scaffold
- PMID: 19670211
- PMCID: PMC2786970
- DOI: 10.1002/pro.218
Alteration of oligomeric state and domain architecture is essential for functional transformation between transferase and hydrolase with the same scaffold
Abstract
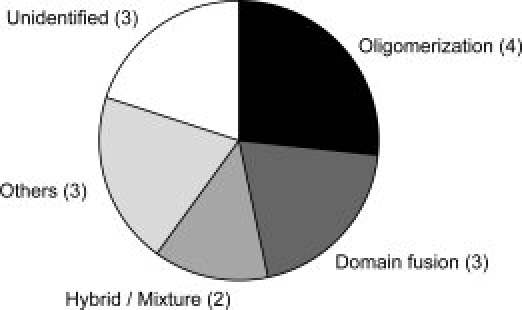
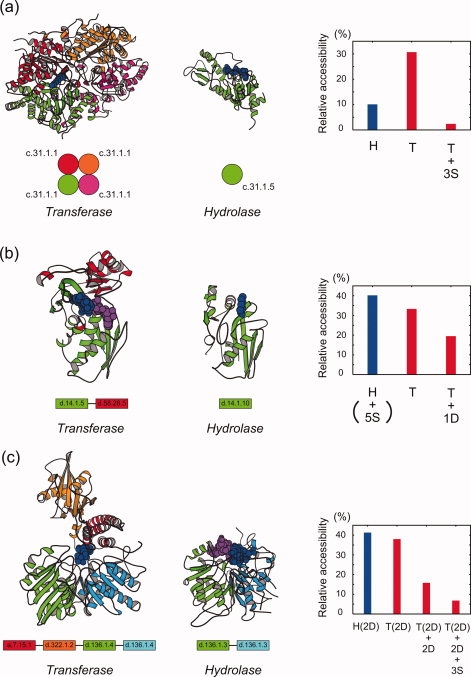
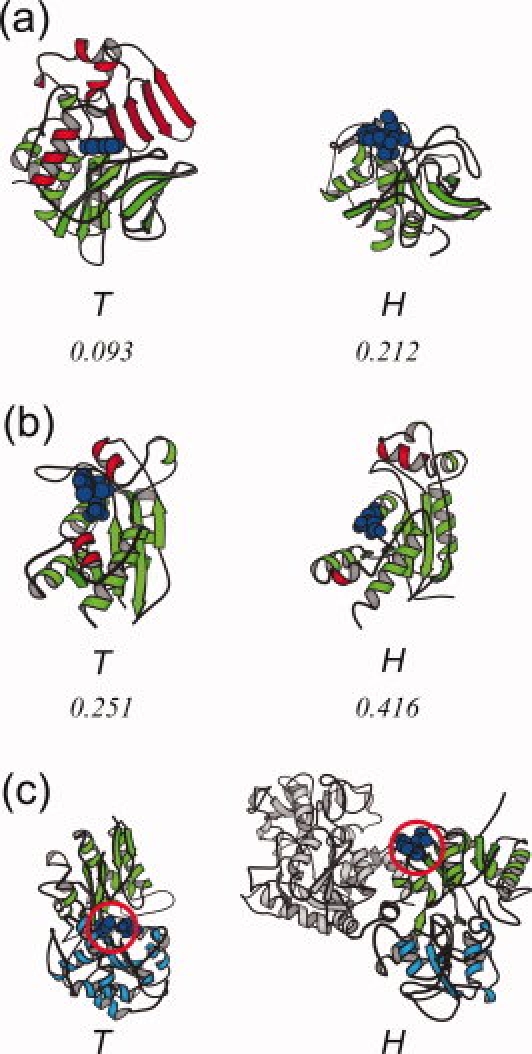
Transferases and hydrolases catalyze different chemical reactions and express different dynamic responses upon ligand binding. To insulate the ligand molecule from the surrounding water, transferases bury it inside the protein by closing the cleft, while hydrolases undergo a small conformational change and leave the ligand molecule exposed to the solvent. Despite these distinct ligand-binding modes, some transferases and hydrolases are homologous. To clarify how such different catalytic modes are possible with the same scaffold, we examined the solvent accessibility of ligand molecules for 15 SCOP superfamilies, each containing both transferase and hydrolase catalytic domains. In contrast to hydrolases, we found that nine superfamilies of transferases use two major strategies, oligomerization and domain fusion, to insulate the ligand molecules. The subunits and domains that were recruited by the transferases often act as a cover for the ligand molecule. The other strategies adopted by transferases to insulate the ligand molecule are the relocation of catalytic sites, the rearrangement of secondary structure elements, and the insertion of peripheral regions. These findings provide insights into how proteins have evolved and acquired distinct functions with a limited number of scaffolds.
Figures
References
-
- Branden C, Tooze J. Introduction to protein structure. 2nd ed. New York: Garland Publishing; 1998.
-
- Glasner ME, Gerlt JA, Babbitt PC. Evolution of enzyme superfamilies. Curr Opin Chem Biol. 2006;10:492–497. - PubMed
-
- Hegyi H, Gerstein M. The relationship between protein structure and function: a comprehensive survey with application to the yeast genome. J Mol Biol. 1999;288:147–164. - PubMed
-
- Martin AC, Orengo CA, Hutchinson EG, Jones S, Karmirantzou M, Laskowski RA, Mitchell JB, Taroni C, Thornton JM. Protein folds and functions. Structure. 1998;6:875–884. - PubMed
-
- Nagano N, Orengo CA, Thornton JM. One fold with many functions: the evolutionary relationships between TIM barrel families based on their sequences, structures and functions. J Mol Biol. 2002;321:741–765. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
