

Absence of direct cyclic nucleotide modulation of mEAG1 and hERG1 channels revealed with fluorescence and electrophysiological methods
- PMID: 19671703
- PMCID: PMC2788851
- DOI: 10.1074/jbc.M109.016337
Absence of direct cyclic nucleotide modulation of mEAG1 and hERG1 channels revealed with fluorescence and electrophysiological methods
Abstract
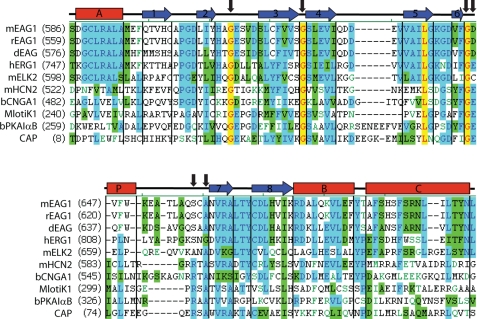
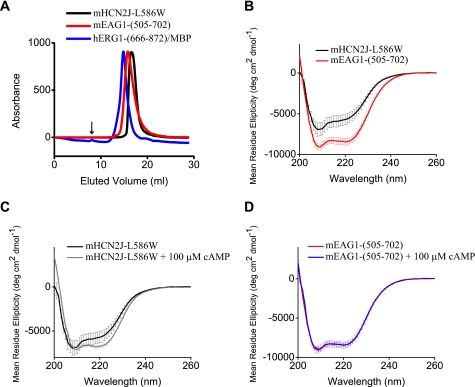
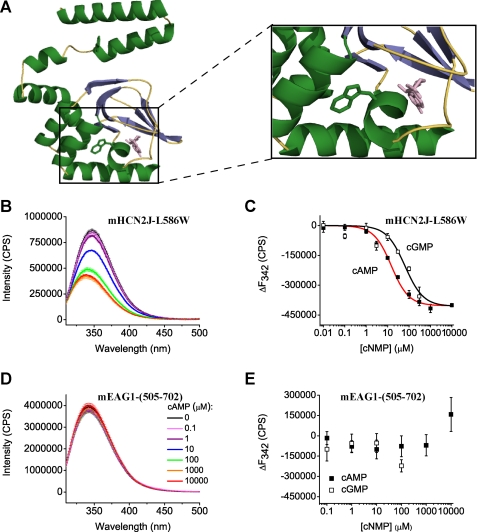
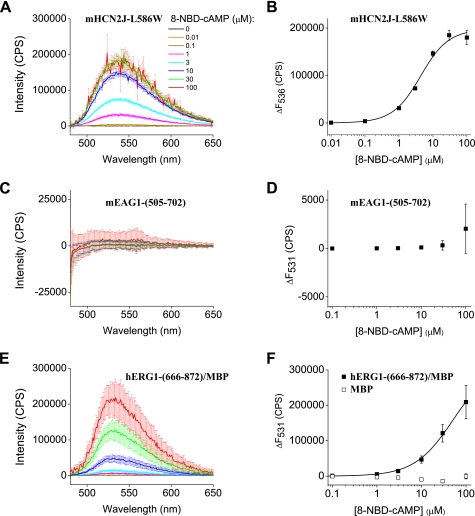
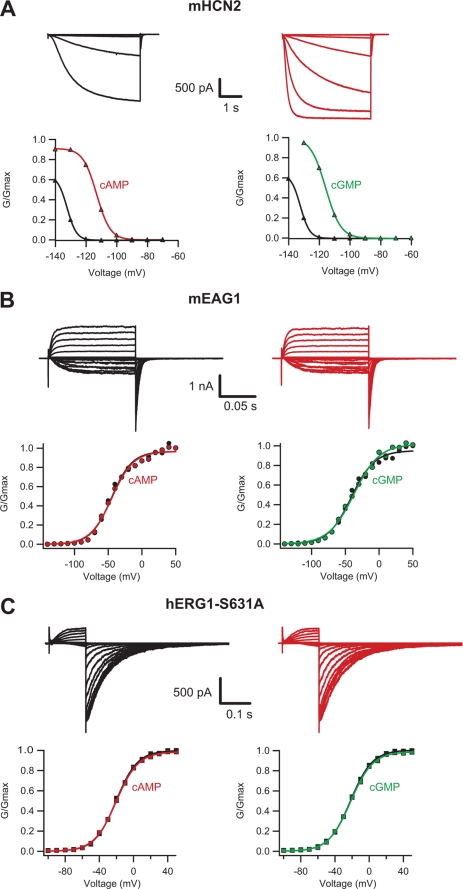
Similar to CNG and HCN channels, EAG and ERG channels contain a cyclic nucleotide binding domain (CNBD) in their C terminus. While cyclic nucleotides have been shown to facilitate opening of CNG and HCN channels, their effect on EAG and ERG channels is less clear. Here we explored cyclic nucleotide binding and modulation of mEAG1 and hERG1 channels with fluorescence and electrophysiology. Binding of cyclic nucleotides to the isolated CNBD of mEAG1 and hERG1 channels was examined with two independent fluorescence-based methods: changes in tryptophan fluorescence and fluorescence of an analog of cAMP, 8-NBD-cAMP. As a positive control for cyclic nucleotide binding we used changes in the fluorescence of the isolated CNBD of mHCN2 channels. Our results indicated that cyclic nucleotides do not bind to the isolated CNBD domain of mEAG1 channels and bind with low affinity (K(d) > or = 51 microm) to the isolated CNBD of hERG1 channels. Consistent with the results on the isolated CNBD, application of cyclic nucleotides to inside-out patches did not affect currents recorded from mEAG1 channels. Surprisingly, despite its low affinity binding to the isolated CNBD, cAMP also had no effect on currents from hERG1 channels even at high concentrations. Our results indicate that cyclic nucleotides do not directly modulate mEAG1 and hERG1 channels. Further studies are necessary to determine if the CNBD in the EAG family of K(+) channels might harbor a binding site for a ligand yet to be uncovered.
Figures
References
-
- Bauer C. K., Schwarz J. R. (2001) J. Membr. Biol. 182, 1–15 - PubMed
-
- Farias L. M., Ocaña D. B., Diaz L., Larrea F., Avila-Chávez E., Cadena A., Hinojosa L. M., Lara G., Villanueva L. A., Vargas C., Hernández-Gallegos E., Camacho-Arroyo I., Dueñas-González A., Pérez-Cárdenas E., Pardo L. A., Morales A., Taja-Chayeb L., Escamilla J., Sánchez-Peña C., Camacho J. (2004) Cancer Res. 64, 6996–7001 - PubMed
-
- Ouadid-Ahidouch H., Le Bourhis X., Roudbaraki M., Toillon R. A., Delcourt P., Prevarskaya N. (2001) Receptors Channels 7, 345–356 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
