

Monocytes promote tumor cell survival in T-cell lymphoproliferative disorders and are impaired in their ability to differentiate into mature dendritic cells
- PMID: 19671921
- PMCID: PMC2756204
- DOI: 10.1182/blood-2009-05-220111
Monocytes promote tumor cell survival in T-cell lymphoproliferative disorders and are impaired in their ability to differentiate into mature dendritic cells
Abstract
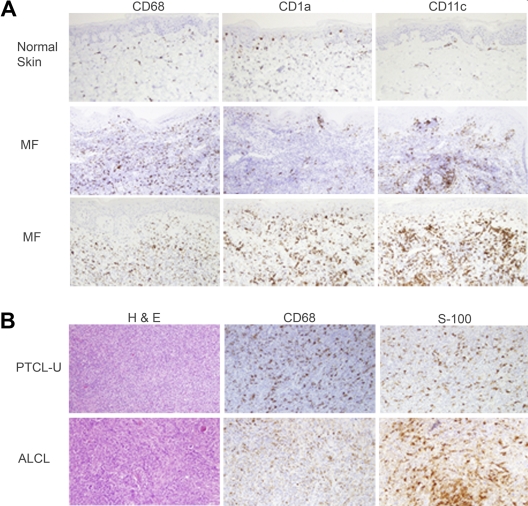
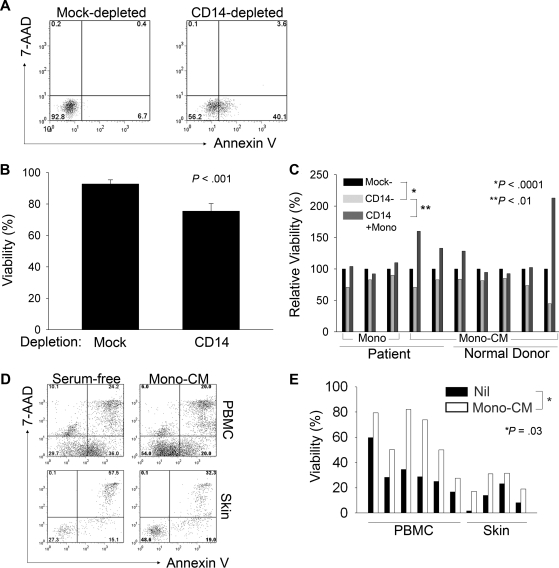
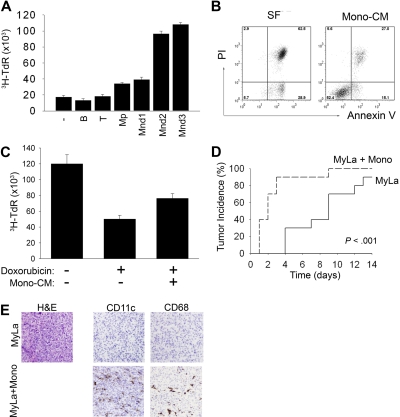
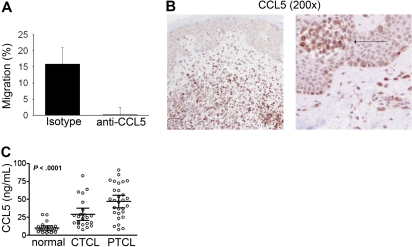
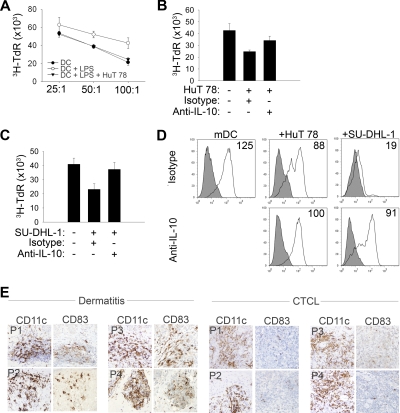
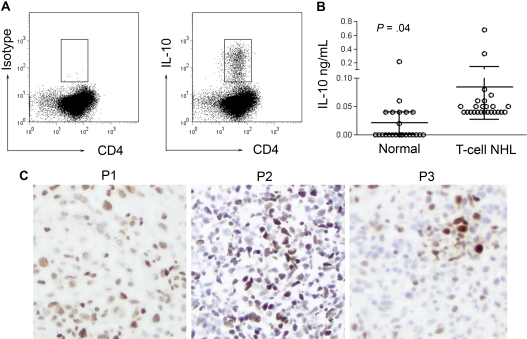
A variety of nonmalignant cells present in the tumor microenvironment promotes tumorigenesis by stimulating tumor cell growth and metastasis or suppressing host immunity. The role of such stromal cells in T-cell lymphoproliferative disorders is incompletely understood. Monocyte-derived cells (MDCs), including professional antigen-presenting cells such as dendritic cells (DCs), play a central role in T-cell biology. Here, we provide evidence that monocytes promote the survival of malignant T cells and demonstrate that MDCs are abundant within the tumor microenvironment of T cell-derived lymphomas. Malignant T cells were observed to remain viable during in vitro culture with autologous monocytes, but cell death was significantly increased after monocyte depletion. Furthermore, monocytes prevent the induction of cell death in T-cell lymphoma lines in response to either serum starvation or doxorubicin, and promote the engraftment of these cells in nonobese diabetic/severe combined immunodeficient mice. Monocytes are actively recruited to the tumor microenvironment by CCL5 (RANTES), where their differentiation into mature DCs is impaired by tumor-derived interleukin-10. Collectively, the data presented demonstrate a previously undescribed role for monocytes in T-cell lymphoproliferative disorders.
Figures
References
-
- Bergers G, Benjamin LE. Tumorigenesis and the angiogenic switch. Nat Rev Cancer. 2003;3(6):401–410. - PubMed
-
- Dave SS, Wright G, Tan B, et al. Prediction of survival in follicular lymphoma based on molecular features of tumor-infiltrating immune cells [see comment]. N Engl J Med. 2004;351(21):2159–2169. - PubMed
-
- Galon J, Costes A, Sanchez-Cabo F, et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome [see comment]. Science. 2006;313(5795):1960–1964. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
