

Vegetally localized Xenopus trim36 regulates cortical rotation and dorsal axis formation
- PMID: 19675128
- PMCID: PMC2730363
- DOI: 10.1242/dev.036855
Vegetally localized Xenopus trim36 regulates cortical rotation and dorsal axis formation
Erratum in
-
Correction: Vegetally localized Xenopus trim36 regulates cortical rotation and dorsal axis formation.Development. 2022 Nov 1;149(21):dev201347. doi: 10.1242/dev.201347. Epub 2022 Oct 24. Development. 2022. PMID: 36278896 Free PMC article. No abstract available.
Abstract
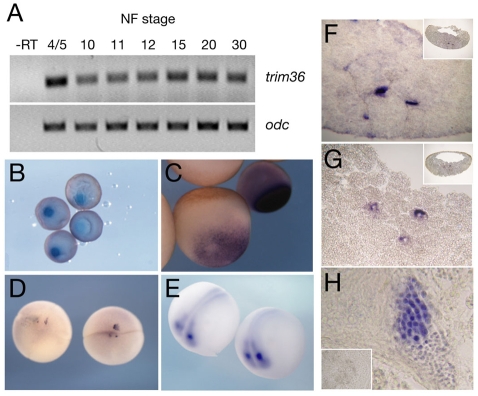
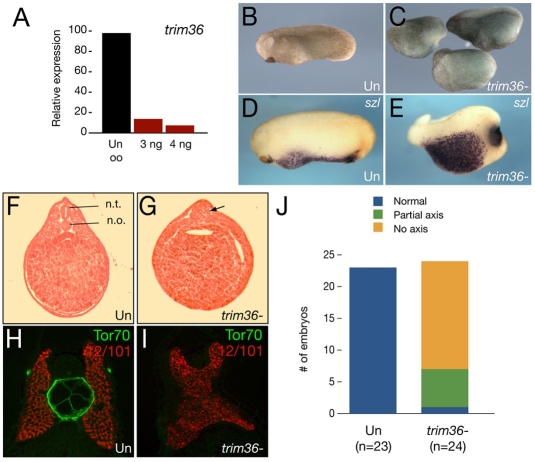
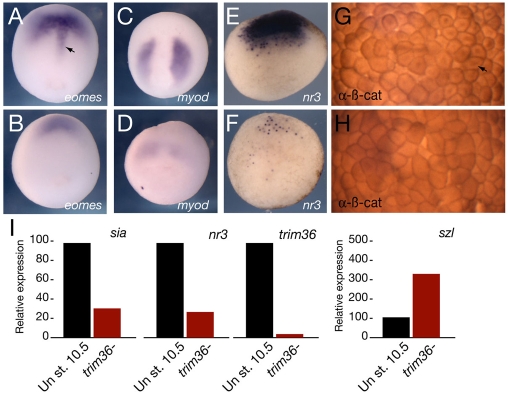
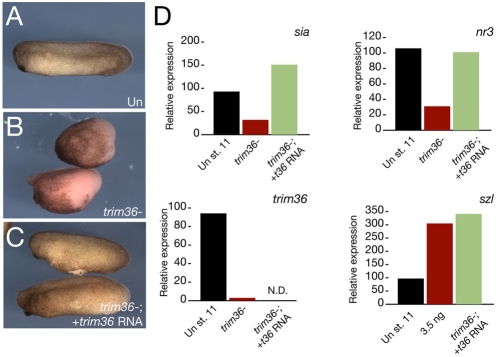
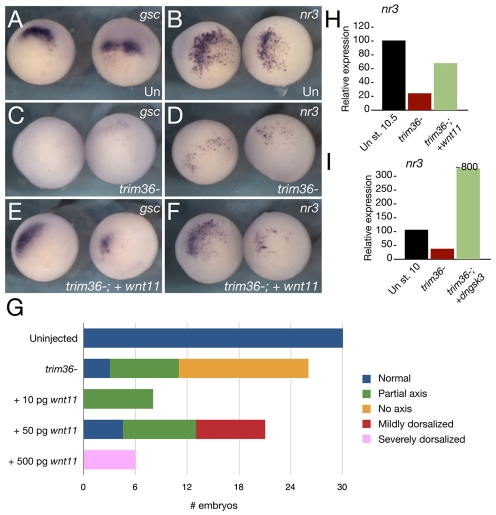
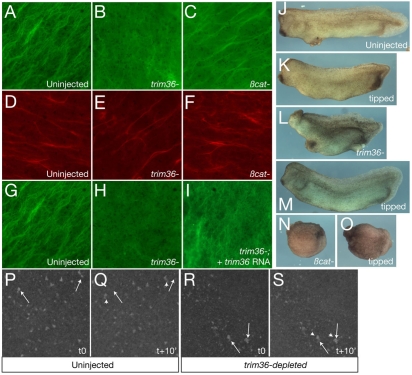
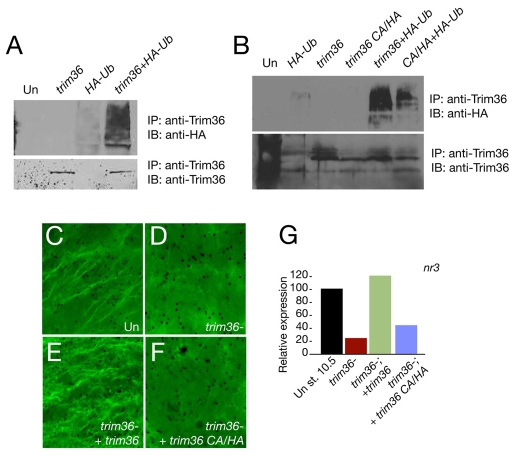
Specification of the dorsoventral axis in Xenopus depends on rearrangements of the egg vegetal cortex following fertilization, concomitant with activation of Wnt/beta-catenin signaling. How these processes are tied together is not clear, but RNAs localized to the vegetal cortex during oogenesis are known to be essential. Despite their importance, few vegetally localized RNAs have been examined in detail. In this study, we describe the identification of a novel localized mRNA, trim36, and characterize its function through maternal loss-of-function experiments. We find that trim36 is expressed in the germ plasm and encodes a ubiquitin ligase of the Tripartite motif-containing (Trim) family. Depletion of maternal trim36 using antisense oligonucleotides results in ventralized embryos and reduced organizer gene expression. We show that injection of wnt11 mRNA rescues this effect, suggesting that Trim36 functions upstream of Wnt/beta-catenin activation. We further find that vegetal microtubule polymerization and cortical rotation are disrupted in trim36-depleted embryos, in a manner dependent on Trim36 ubiquitin ligase activity. Additionally, these embryos can be rescued by tipping the eggs 90 degrees relative to the animal-vegetal axis. Taken together, our results suggest a role for Trim36 in controlling the stability of proteins regulating microtubule polymerization during cortical rotation, and subsequently axis formation.
Figures
Similar articles
-
Maternal Dead-End1 is required for vegetal cortical microtubule assembly during Xenopus axis specification.Development. 2013 Jun;140(11):2334-44. doi: 10.1242/dev.094748. Epub 2013 Apr 24. Development. 2013. PMID: 23615278 Free PMC article.
-
The dynamics of plus end polarization and microtubule assembly during Xenopus cortical rotation.Dev Biol. 2015 May 15;401(2):249-63. doi: 10.1016/j.ydbio.2015.01.028. Epub 2015 Mar 7. Dev Biol. 2015. PMID: 25753733 Free PMC article.
-
Dorsal determinants in the Xenopus egg are firmly associated with the vegetal cortex and behave like activators of the Wnt pathway.Dev Biol. 1997 Nov 1;191(1):69-79. doi: 10.1006/dbio.1997.8710. Dev Biol. 1997. PMID: 9356172
-
Cortical rotation and messenger RNA localization in Xenopus axis formation.Wiley Interdiscip Rev Dev Biol. 2012 May-Jun;1(3):371-88. doi: 10.1002/wdev.29. Epub 2012 Jan 19. Wiley Interdiscip Rev Dev Biol. 2012. PMID: 23801488 Review.
-
Move it or lose it: axis specification in Xenopus.Development. 2004 Aug;131(15):3491-9. doi: 10.1242/dev.01284. Development. 2004. PMID: 15262887 Review.
Cited by
-
Characterization and functional roles of paternal RNAs in 2-4 cell bovine embryos.Sci Rep. 2019 Dec 30;9(1):20347. doi: 10.1038/s41598-019-55868-3. Sci Rep. 2019. PMID: 31889064 Free PMC article.
-
Lysosomal degradation of the maternal dorsal determinant Hwa safeguards dorsal body axis formation.EMBO Rep. 2021 Dec 6;22(12):e53185. doi: 10.15252/embr.202153185. Epub 2021 Oct 15. EMBO Rep. 2021. PMID: 34652064 Free PMC article.
-
Germ plasm in Eleutherodactylus coqui, a direct developing frog with large eggs.Evodevo. 2011 Oct 6;2:20. doi: 10.1186/2041-9139-2-20. Evodevo. 2011. PMID: 21978790 Free PMC article.
-
Identification of germ plasm-associated transcripts by microarray analysis of Xenopus vegetal cortex RNA.Dev Dyn. 2010 Jun;239(6):1838-48. doi: 10.1002/dvdy.22304. Dev Dyn. 2010. PMID: 20503379 Free PMC article.
-
Vertebrate Axial Patterning: From Egg to Asymmetry.Adv Exp Med Biol. 2017;953:209-306. doi: 10.1007/978-3-319-46095-6_6. Adv Exp Med Biol. 2017. PMID: 27975274 Free PMC article. Review.
References
-
- Belenkaya, T. Y., Han, C., Standley, H. J., Lin, X., Houston, D. W., Heasman, J. and Lin, X. (2002). pygopus Encodes a nuclear protein essential for wingless/Wnt signaling. Development 129, 4089-4101. - PubMed
-
- Brown, E. E., Margelot, K. M. and Danilchik, M. V. (1994). Provisional bilateral symmetry in Xenopus eggs is established during maturation. Zygote 2, 213-220. - PubMed
-
- Collavin, L. and Kirschner, M. W. (2003). The secreted Frizzled-related protein Sizzled functions as a negative feedback regulator of extreme ventral mesoderm. Development 130, 805-816. - PubMed
-
- Darras, S., Marikawa, Y., Elinson, R. P. and Lemaire, P. (1997). Animal and vegetal pole cells of early Xenopus embryos respond differently to maternal dorsal determinants: implications for the patterning of the organiser. Development 124, 4275-4286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
