

Modulation of single-cell IgG secretion frequency and rates in human memory B cells by CpG DNA, CD40L, IL-21, and cell division
- PMID: 19675172
- PMCID: PMC2765874
- DOI: 10.4049/jimmunol.0804233
Modulation of single-cell IgG secretion frequency and rates in human memory B cells by CpG DNA, CD40L, IL-21, and cell division
Abstract
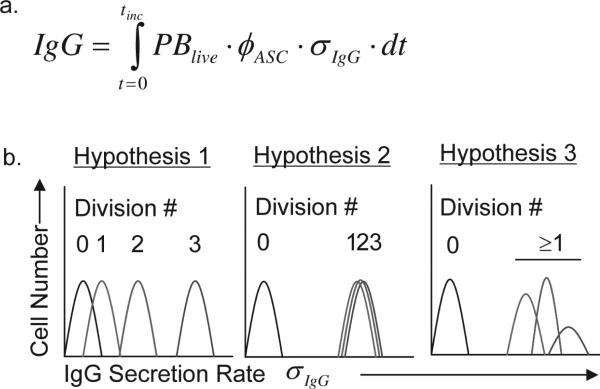
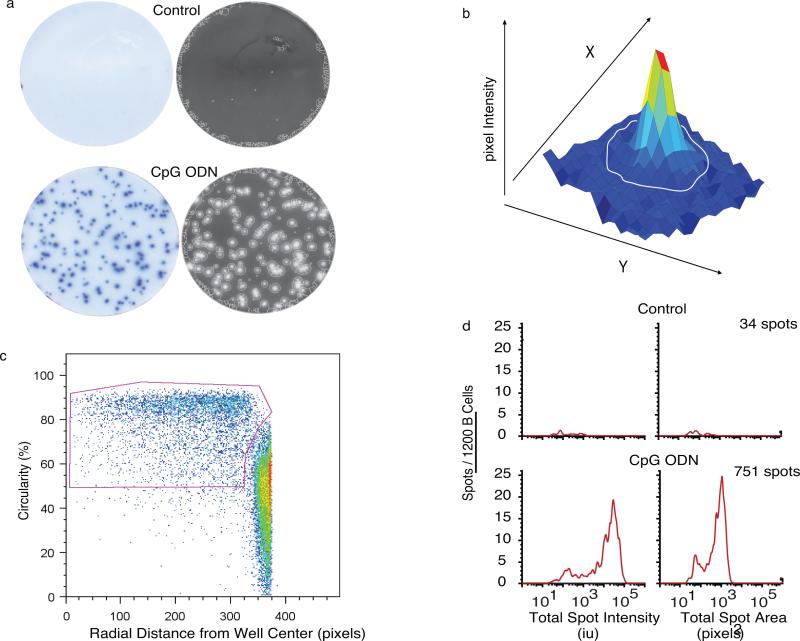
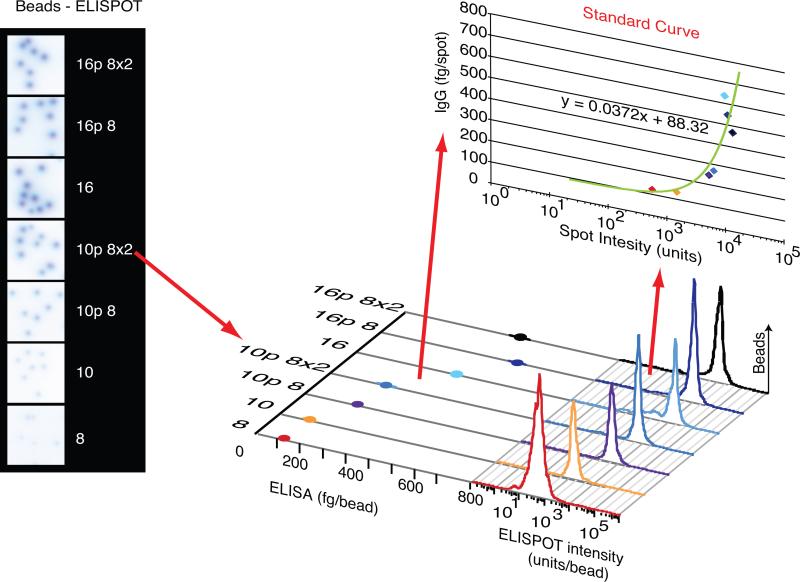
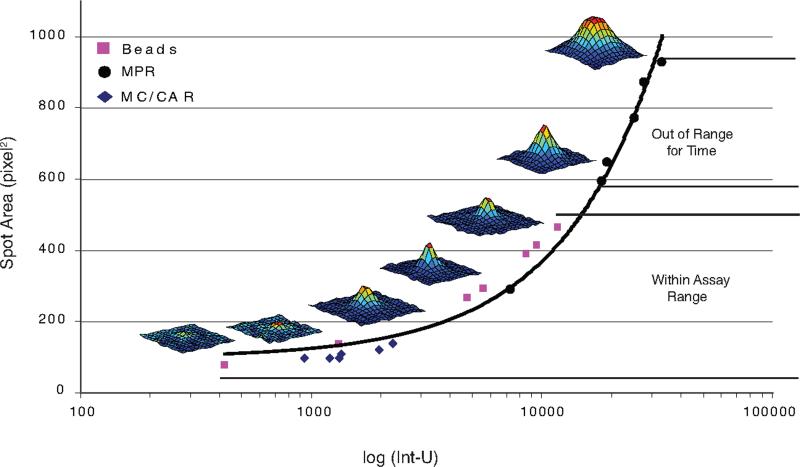
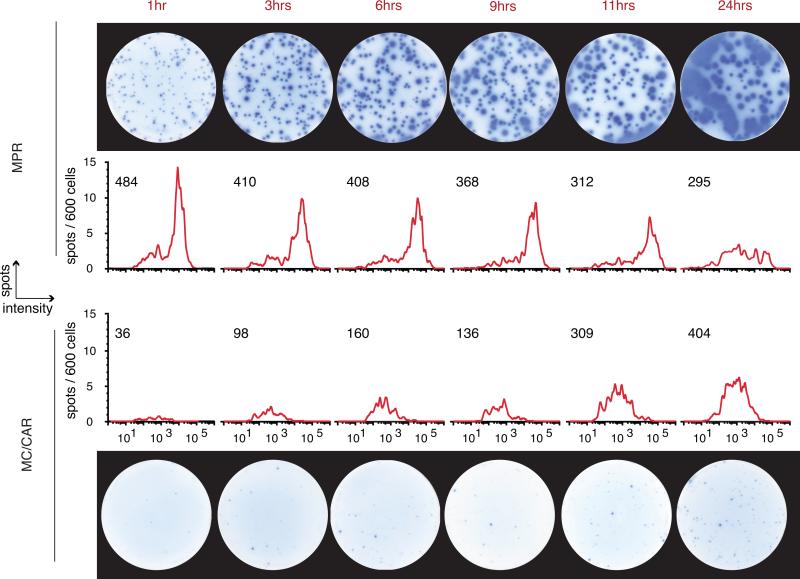
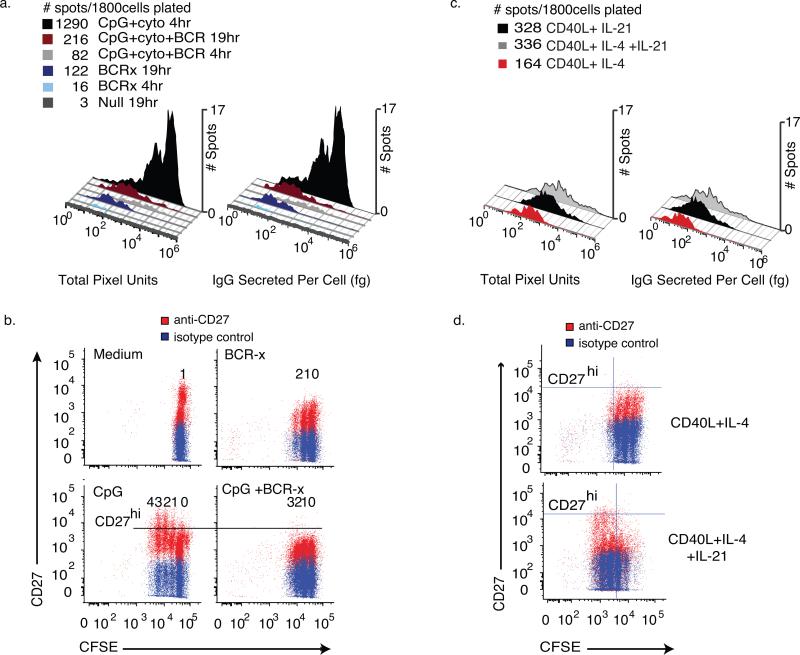
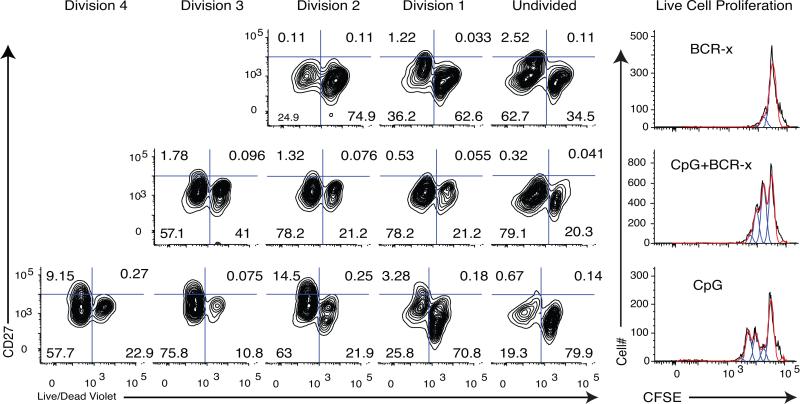
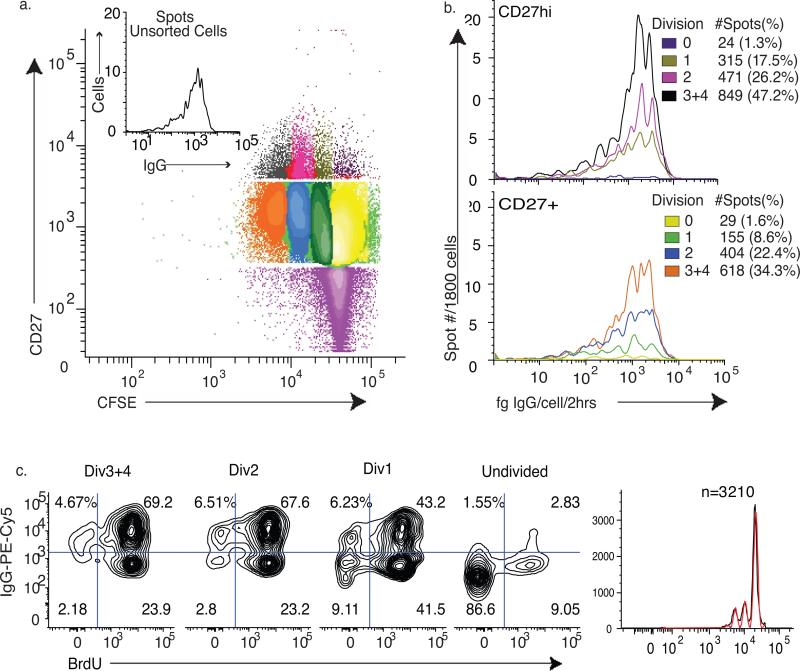
During the recall response by CD27(+) IgG class-switched human memory B cells, total IgG secreted is a function of the following: 1) the number of IgG-secreting cells (IgG-SC), and 2) the secretion rate of each cell. In this study, we report the quantitative ELISPOT method for simultaneous estimation of single-cell IgG secretion rates and secreting cell frequencies in human B cell populations. We found that CD27(+) IgM(-) memory B cells activated with CpG and cytokines had considerable heterogeneity in the IgG secretion rates, with two major secretion rate subpopulations. BCR cross-linking reduced the frequency of cells with high per-cell IgG secretion rates, with a parallel decrease in CD27(high) B cell blasts. Increased cell death may account for the BCR-stimulated reduction in high-rate IgG-SC CD27(high) B cell blasts. In contrast, the addition of IL-21 to CD40L plus IL-4-activated human memory B cells induced a high-rate IgG-SC population in B cells with otherwise low per-cell IgG secretion rates. The profiles of human B cell IgG secretion rates followed the same biphasic distribution and range irrespective of division class. This, along with the presence of non-IgG-producing, dividing B cells in CpG plus cytokine-activated B memory B cell populations, is suggestive of an on/off switch regulating IgG secretion. Finally, these data support a mixture model of IgG secretion in which IgG secreted over time is modulated by the frequency of IgG-SC and the distribution of their IgG secretion rates.
Figures
Similar articles
-
IL-12p70-dependent Th1 induction by human B cells requires combined activation with CD40 ligand and CpG DNA.J Immunol. 2004 Jan 15;172(2):954-63. doi: 10.4049/jimmunol.172.2.954. J Immunol. 2004. PMID: 14707068
-
Peripheral blood CD27+ IgG+ B cells rapidly proliferate and differentiate into immunoglobulin-secreting cells after exposure to low CD154 interaction.Immunology. 2009 Sep;128(1 Suppl):e353-65. doi: 10.1111/j.1365-2567.2008.02976.x. Epub 2008 Nov 7. Immunology. 2009. PMID: 19016905 Free PMC article.
-
A division-linked mechanism for the rapid generation of Ig-secreting cells from human memory B cells.J Immunol. 2003 Jan 1;170(1):261-9. doi: 10.4049/jimmunol.170.1.261. J Immunol. 2003. PMID: 12496408
-
CD40-dependent and -independent activation of human tonsil B cells by CpG oligodeoxynucleotides.Eur J Immunol. 2003 Jun;33(6):1576-85. doi: 10.1002/eji.200323444. Eur J Immunol. 2003. PMID: 12778475
-
An optimized assay for the enumeration of antigen-specific memory B cells in different compartments of the human body.J Immunol Methods. 2010 Jun 30;358(1-2):56-65. doi: 10.1016/j.jim.2010.03.009. Epub 2010 Mar 17. J Immunol Methods. 2010. PMID: 20302874
Cited by
-
Nanowell-based immunoassays for measuring single-cell secretion: characterization of transport and surface binding.Anal Chem. 2014 Dec 2;86(23):11562-9. doi: 10.1021/ac4030297. Epub 2014 Nov 17. Anal Chem. 2014. PMID: 25347613 Free PMC article.
-
Optimisation of ex vivo memory B cell expansion/differentiation for interrogation of rare peripheral memory B cell subset responses.Wellcome Open Res. 2017 Oct 5;2:97. doi: 10.12688/wellcomeopenres.11386.2. eCollection 2017. Wellcome Open Res. 2017. PMID: 29588920 Free PMC article.
-
The Role of γδ T Cells in Systemic Lupus Erythematosus.J Immunol Res. 2016;2016:2932531. doi: 10.1155/2016/2932531. Epub 2016 Feb 11. J Immunol Res. 2016. PMID: 26981547 Free PMC article. Review.
-
A Label-free Technique for the Spatio-temporal Imaging of Single Cell Secretions.J Vis Exp. 2015 Nov 23;(105):53120. doi: 10.3791/53120. J Vis Exp. 2015. PMID: 26650542 Free PMC article.
-
Hyperactivation and in situ recruitment of inflammatory Vδ2 T cells contributes to disease pathogenesis in systemic lupus erythematosus.Sci Rep. 2015 Sep 23;5:14432. doi: 10.1038/srep14432. Sci Rep. 2015. PMID: 26395317 Free PMC article.
References
-
- Tangye SG, Avery DT, Hodgkin PD. A division-linked mechanism for the rapid generation of Ig-secreting cells from human memory B cells. J Immunol. 2003;170:261–269. - PubMed
-
- Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H, Matsumoto M, Hoshino K, Wagner H, Takeda K, Akira S. A Toll-like receptor recognizes bacterial DNA. Nature. 2000;408:740–745. - PubMed
-
- Bernasconi NL, Traggiai E, Lanzavecchia A. Maintenance of serological memory by polyclonal activation of human memory B cells. Science. 2002;298:2199–2202. - PubMed
-
- Shiao RT, McLeskey SB, Khera SY, Wolfson A, Freter CE. Mechanisms of inhibition of IL-6-mediated immunoglobulin secretion by dexamethasone and suramin in human lymphoid and myeloma cell lines. Leukemia & lymphoma. 1996;21:293–303. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
