

Enhanced calcium buffering in F344 rat cholinergic basal forebrain neurons is associated with age-related cognitive impairment
- PMID: 19675291
- PMCID: PMC2775378
- DOI: 10.1152/jn.00301.2009
Enhanced calcium buffering in F344 rat cholinergic basal forebrain neurons is associated with age-related cognitive impairment
Abstract
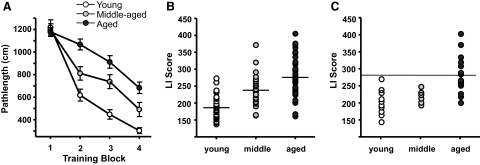
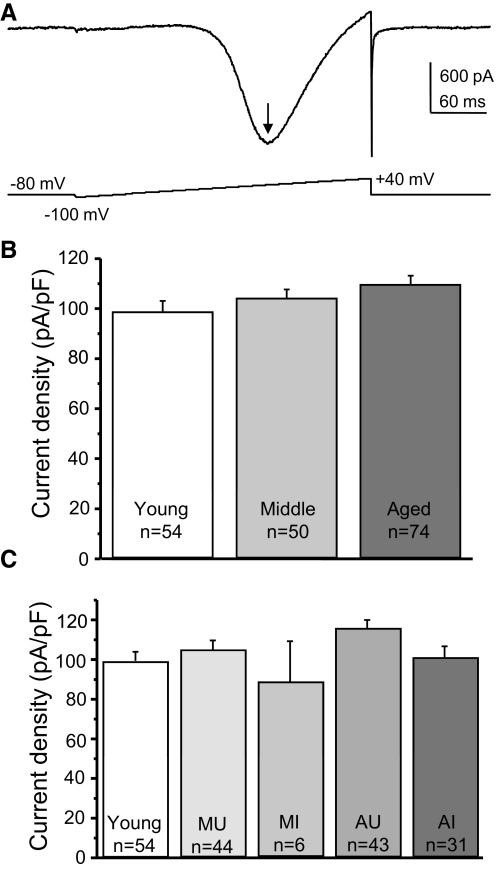
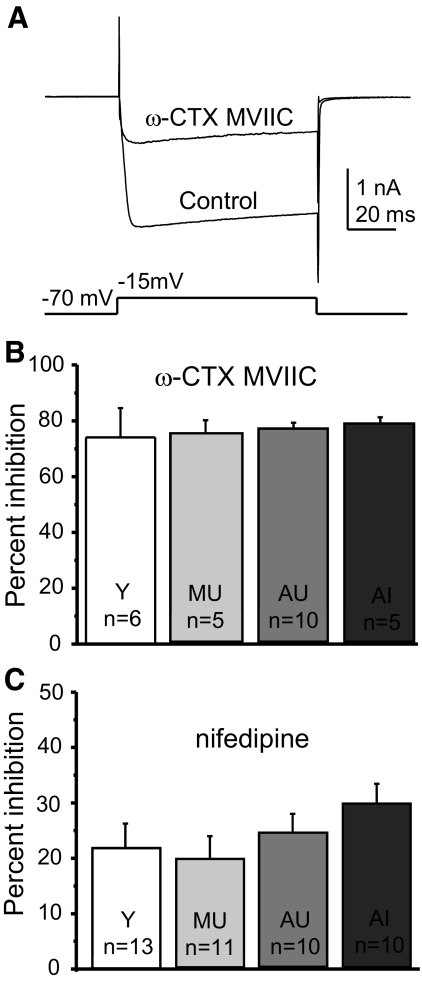
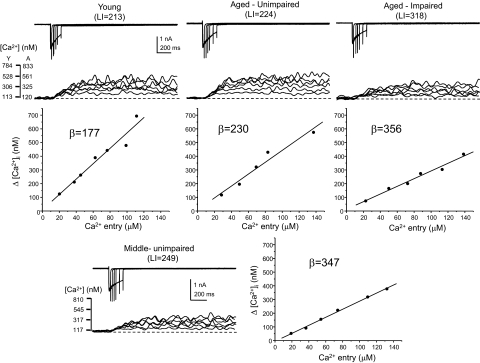
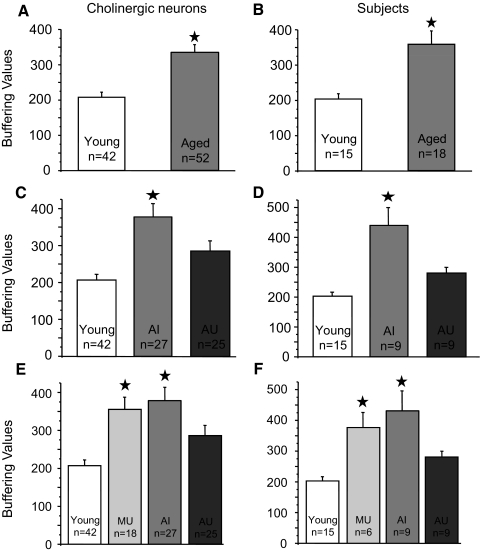
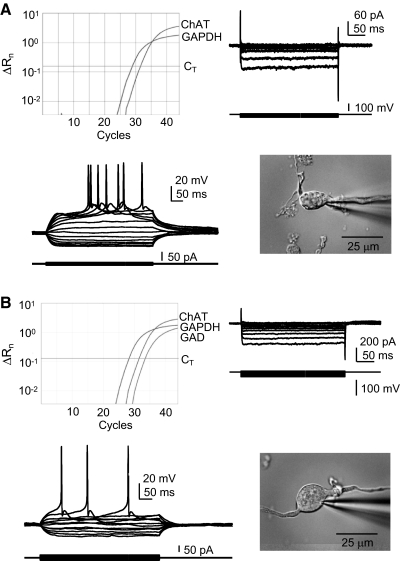
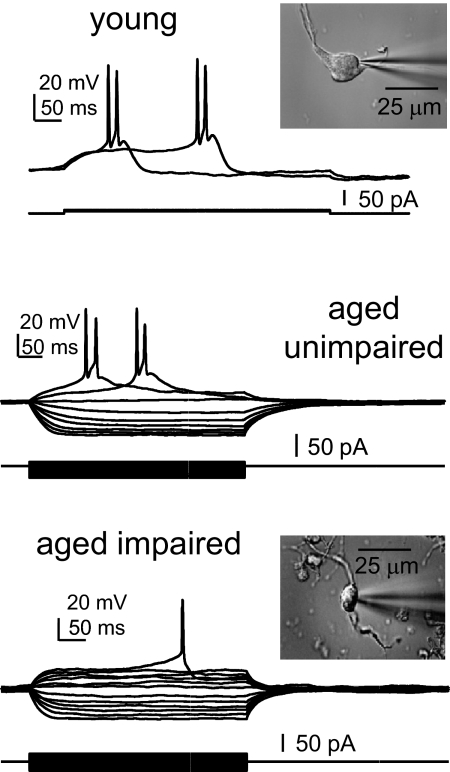
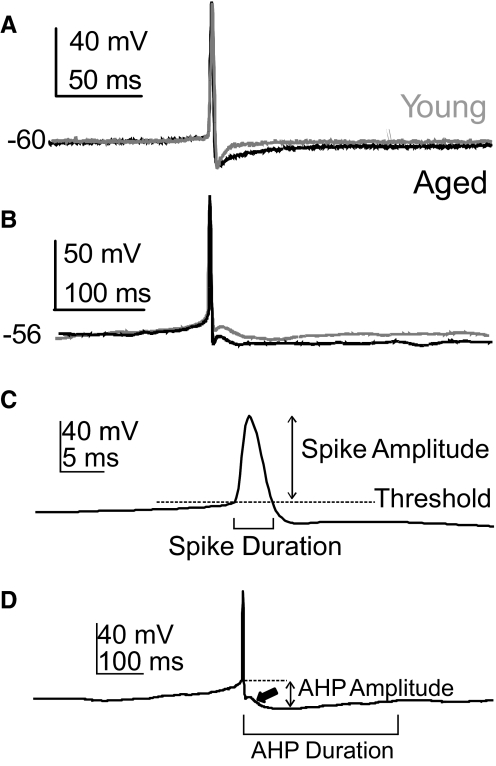
Alterations in neuronal Ca(2+) homeostasis are important determinants of age-related cognitive impairment. We examined the Ca(2+) influx, buffering, and electrophysiology of basal forebrain neurons in adult, middle-aged, and aged male F344 behaviorally assessed rats. Middle-aged and aged rats were characterized as cognitively impaired or unimpaired by water maze performance relative to young cohorts. Patch-clamp experiments were conducted on neurons acutely dissociated from medial septum/nucleus of the diagonal band with post hoc identification of phenotypic marker mRNA using single-cell RT-PCR. We measured whole cell calcium and barium currents and dissected these currents using pharmacological agents. We combined Ca(2+) current recording with Ca(2+)-sensitive ratiometric microfluorimetry to measure Ca(2+) buffering. Additionally, we sought changes in neuronal firing properties using current-clamp recording. There were no age- or cognition-related changes in the amplitudes or fractional compositions of the whole cell Ca(2+) channel currents. However, Ca(2+) buffering was significantly enhanced in cholinergic neurons from aged cognitively impaired rats. Moreover, increased Ca(2+) buffering was present in middle-aged rats that were not cognitively impaired. Firing properties were largely unchanged with age or cognitive status, except for an increase in the slow afterhyperpolarization in aged cholinergic neurons, independent of cognitive status. Furthermore, acutely dissociated basal forebrain neurons in which choline acetyltransferase mRNA was detected had the electrophysiological profiles of identified cholinergic neurons. We conclude that enhanced Ca(2+) buffering by cholinergic basal forebrain neurons may be important during aging.
Figures
Similar articles
-
Characterization of age-related changes in synaptic transmission onto F344 rat basal forebrain cholinergic neurons using a reduced synaptic preparation.J Neurophysiol. 2014 Jan;111(2):273-86. doi: 10.1152/jn.00129.2013. Epub 2013 Oct 16. J Neurophysiol. 2014. PMID: 24133226 Free PMC article.
-
Low voltage-activated calcium and fast tetrodotoxin-resistant sodium currents define subtypes of cholinergic and noncholinergic neurons in rat basal forebrain.Brain Res Mol Brain Res. 2005 Apr 4;134(2):226-38. doi: 10.1016/j.molbrainres.2004.10.041. Epub 2005 Jan 21. Brain Res Mol Brain Res. 2005. PMID: 15836920
-
Increased calcium buffering in basal forebrain neurons during aging.J Neurophysiol. 1998 Jul;80(1):350-64. doi: 10.1152/jn.1998.80.1.350. J Neurophysiol. 1998. PMID: 9658056
-
Calcium buffering systems and calcium signaling in aged rat basal forebrain neurons.Aging Cell. 2007 Jun;6(3):297-305. doi: 10.1111/j.1474-9726.2007.00293.x. Aging Cell. 2007. PMID: 17517040 Free PMC article. Review.
-
Immunolesion by 192IgG-saporin of rat basal forebrain cholinergic system: a useful tool to produce cortical cholinergic dysfunction.Prog Brain Res. 1996;109:253-64. doi: 10.1016/s0079-6123(08)62109-3. Prog Brain Res. 1996. PMID: 9009714 Review.
Cited by
-
Calcium's role as nuanced modulator of cellular physiology in the brain.Biochem Biophys Res Commun. 2017 Feb 19;483(4):981-987. doi: 10.1016/j.bbrc.2016.08.105. Epub 2016 Aug 20. Biochem Biophys Res Commun. 2017. PMID: 27553276 Free PMC article. Review.
-
Glial Cells Response in Stroke.Cell Mol Neurobiol. 2023 Jan;43(1):99-113. doi: 10.1007/s10571-021-01183-3. Epub 2022 Jan 23. Cell Mol Neurobiol. 2023. PMID: 35066715 Free PMC article. Review.
-
Susceptibility to Calcium Dysregulation during Brain Aging.Front Aging Neurosci. 2009 Nov 27;1:2. doi: 10.3389/neuro.24.002.2009. eCollection 2009. Front Aging Neurosci. 2009. PMID: 20552053 Free PMC article.
-
Effect of a water-maze procedure on the redox mechanisms in brain parts of aged rats.Front Aging Neurosci. 2015 Mar 11;7:29. doi: 10.3389/fnagi.2015.00029. eCollection 2015. Front Aging Neurosci. 2015. PMID: 25814952 Free PMC article.
-
Electrophysiological and Imaging Calcium Biomarkers of Aging in Male and Female 5×FAD Mice.J Alzheimers Dis. 2020;78(4):1419-1438. doi: 10.3233/JAD-200109. J Alzheimers Dis. 2020. PMID: 33164928 Free PMC article.
References
-
- Bartus RT. On neurodegenerative diseases, models, and treatment strategies: lessons learned and lessons forgotten a generation following the cholinergic hypothesis. Exp Neurol 163: 495–529, 2000 - PubMed
-
- Baxter MG, Frick KM, Price DL, Breckler SJ, Markowska AL, Gorman LK. Presynaptic markers of cholinergic function in the rat brain: relationship with age and cognitive status. Neuroscience 89: 771–780, 1999 - PubMed
-
- Berridge MJ, Lipp P, Bootman MD. The versatility and universality of calcium signalling. Nat Rev Mol Cell Biol 1: 11–21, 2000 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous