

Selective interaction of syntaxin 1A with KCNQ2: possible implications for specific modulation of presynaptic activity
- PMID: 19675672
- PMCID: PMC2721677
- DOI: 10.1371/journal.pone.0006586
Selective interaction of syntaxin 1A with KCNQ2: possible implications for specific modulation of presynaptic activity
Abstract
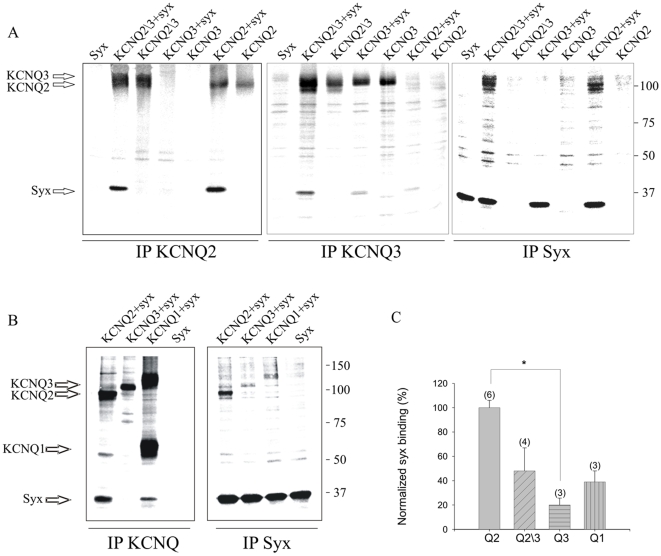
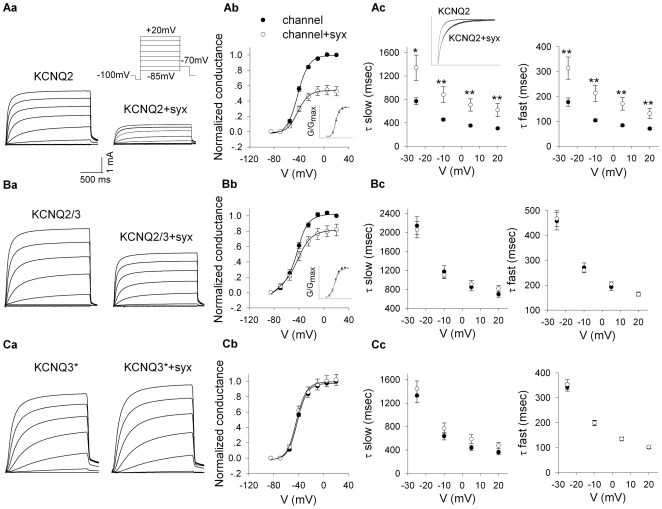
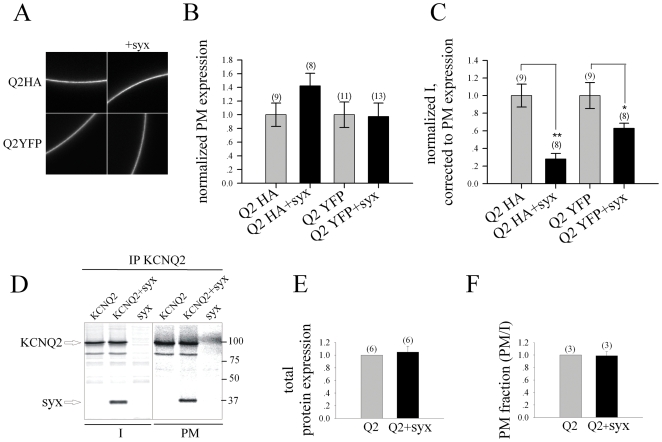
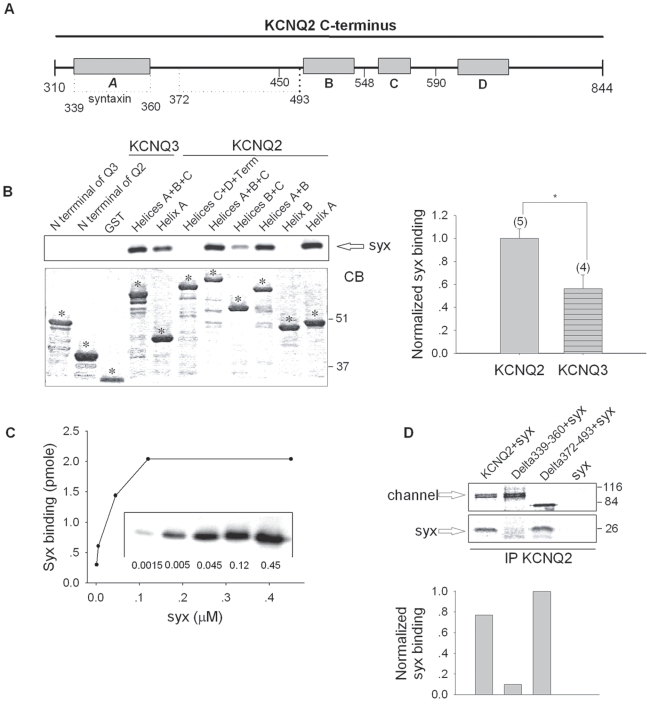
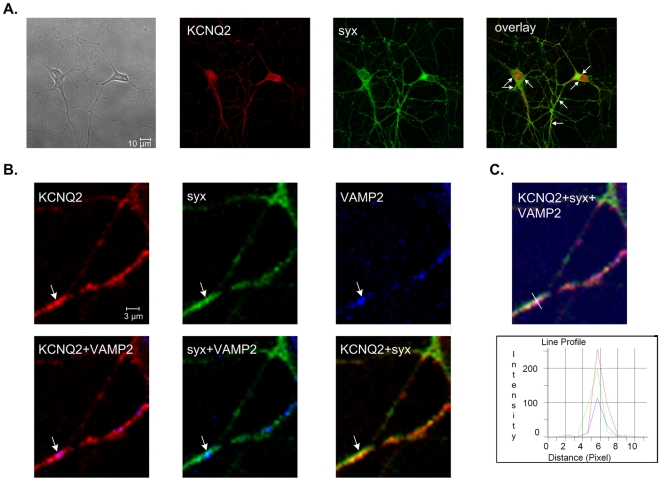
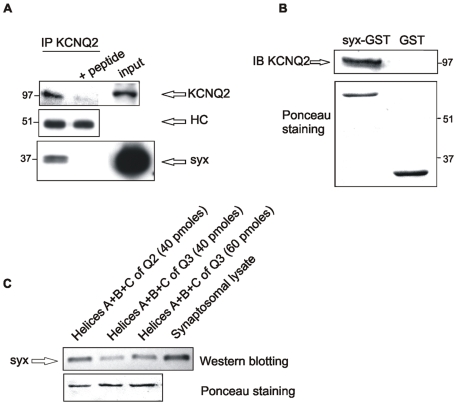
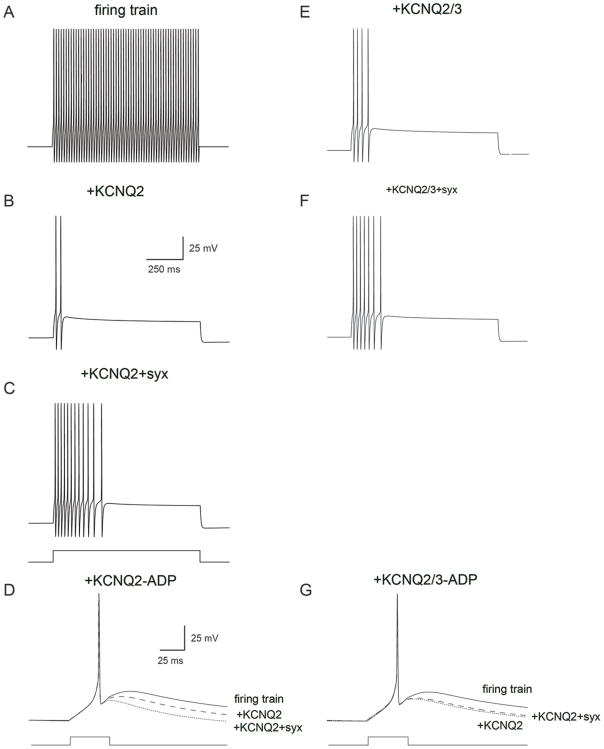
KCNQ2/KCNQ3 channels are the molecular correlates of the neuronal M-channels, which play a major role in the control of neuronal excitability. Notably, they differ from homomeric KCNQ2 channels in their distribution pattern within neurons, with unique expression of KCNQ2 in axons and nerve terminals. Here, combined reciprocal coimmunoprecipitation and two-electrode voltage clamp analyses in Xenopus oocytes revealed a strong association of syntaxin 1A, a major component of the exocytotic SNARE complex, with KCNQ2 homomeric channels resulting in a approximately 2-fold reduction in macroscopic conductance and approximately 2-fold slower activation kinetics. Remarkably, the interaction of KCNQ2/Q3 heteromeric channels with syntaxin 1A was significantly weaker and KCNQ3 homomeric channels were practically resistant to syntaxin 1A. Analysis of different KCNQ2 and KCNQ3 chimeras and deletion mutants combined with in-vitro binding analysis pinpointed a crucial C-terminal syntaxin 1A-association domain in KCNQ2. Pull-down and coimmunoprecipitation analyses in hippocampal and cortical synaptosomes demonstrated a physical interaction of brain KCNQ2 with syntaxin 1A, and confocal immunofluorescence microscopy showed high colocalization of KCNQ2 and syntaxin 1A at presynaptic varicosities. The selective interaction of syntaxin 1A with KCNQ2, combined with a numerical simulation of syntaxin 1A's impact in a firing-neuron model, suggest that syntaxin 1A's interaction is targeted at regulating KCNQ2 channels to fine-tune presynaptic transmitter release, without interfering with the function of KCNQ2/3 channels in neuronal firing frequency adaptation.
Conflict of interest statement
Figures
Similar articles
-
Regulation of neuronal M-channel gating in an isoform-specific manner: functional interplay between calmodulin and syntaxin 1A.J Neurosci. 2011 Oct 5;31(40):14158-71. doi: 10.1523/JNEUROSCI.2666-11.2011. J Neurosci. 2011. PMID: 21976501 Free PMC article.
-
A possible link between KCNQ2- and STXBP1-related encephalopathies: STXBP1 reduces the inhibitory impact of syntaxin-1A on M current.Epilepsia. 2017 Dec;58(12):2073-2084. doi: 10.1111/epi.13927. Epub 2017 Oct 25. Epilepsia. 2017. PMID: 29067685
-
Structural determinants of M-type KCNQ (Kv7) K+ channel assembly.J Neurosci. 2006 Apr 5;26(14):3757-66. doi: 10.1523/JNEUROSCI.5017-05.2006. J Neurosci. 2006. PMID: 16597729 Free PMC article.
-
Direct interaction of target SNAREs with the Kv2.1 channel. Modal regulation of channel activation and inactivation gating.J Biol Chem. 2003 Sep 5;278(36):34320-30. doi: 10.1074/jbc.M304943200. Epub 2003 Jun 13. J Biol Chem. 2003. PMID: 12807875
-
Novel KCNQ2 and KCNQ3 mutations in a large cohort of families with benign neonatal epilepsy: first evidence for an altered channel regulation by syntaxin-1A.Hum Mutat. 2014 Mar;35(3):356-67. doi: 10.1002/humu.22500. Epub 2014 Jan 13. Hum Mutat. 2014. PMID: 24375629
Cited by
-
Role of neuronal potassium M-channels in sympathetic regulation of cardiac function.J Physiol. 2011 Jun 1;589(Pt 11):2659-60. doi: 10.1113/jphysiol.2011.210161. J Physiol. 2011. PMID: 21632525 Free PMC article. No abstract available.
-
Regulation of neuronal M-channel gating in an isoform-specific manner: functional interplay between calmodulin and syntaxin 1A.J Neurosci. 2011 Oct 5;31(40):14158-71. doi: 10.1523/JNEUROSCI.2666-11.2011. J Neurosci. 2011. PMID: 21976501 Free PMC article.
-
Sub- and suprathreshold adaptation currents have opposite effects on frequency tuning.J Physiol. 2012 Oct 1;590(19):4839-58. doi: 10.1113/jphysiol.2012.234401. Epub 2012 Jun 25. J Physiol. 2012. PMID: 22733663 Free PMC article.
-
The Role of Kv7.2 in Neurodevelopment: Insights and Gaps in Our Understanding.Front Physiol. 2020 Oct 28;11:570588. doi: 10.3389/fphys.2020.570588. eCollection 2020. Front Physiol. 2020. PMID: 33192566 Free PMC article. Review.
-
The Role of Kv7 Channels in Neural Plasticity and Behavior.Front Physiol. 2020 Sep 18;11:568667. doi: 10.3389/fphys.2020.568667. eCollection 2020. Front Physiol. 2020. PMID: 33071824 Free PMC article. Review.
References
-
- Brown D. M-currents: an update. Trends Neurosci. 1988;11:294–299. - PubMed
-
- Marrion NV. Control of M-current. Annu Rev Physiol. 1997;59:483–504. - PubMed
-
- Jentsch TJ. Neuronal KCNQ potassium channels: physiology and role in disease. Nat Rev Neurosci. 2000;1:21–30. - PubMed
-
- Rogawski MA. KCNQ2/KCNQ3 K+ channels and the molecular pathogenesis of epilepsy: implications for therapy. Trends Neurosci. 2000;23:393–398. - PubMed
-
- Wang HS, Pan Z, Shi W, Brown BS, Wymore RS, et al. KCNQ2 and KCNQ3 potassium channel subunits: molecular correlates of the M-channel. Science. 1998;282:1890–1893. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
