

TRPC channel-mediated neuroprotection by PDGF involves Pyk2/ERK/CREB pathway
- PMID: 19680266
- PMCID: PMC2783976
- DOI: 10.1038/cdd.2009.108
TRPC channel-mediated neuroprotection by PDGF involves Pyk2/ERK/CREB pathway
Abstract
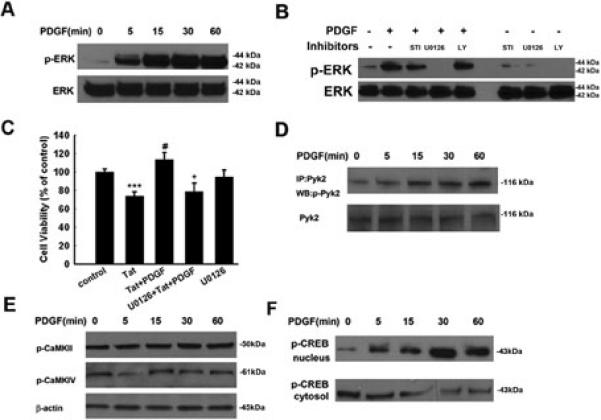
Platelet-derived growth factor-BB (PDGF) has been reported to provide tropic support for neurons in the central nervous system. The protective role of PDGF on dopaminergic neurons, especially in the context of HIV-associated dementia (HAD), however, remains largely unknown. Here, we show that exogenous PDGF was neuroprotective against toxicity induced by HIV-1 Tat in primary midbrain neurons. Furthermore, we report the involvement of transient receptor potential canonical (TRPC) channels in PDGF-mediated neuroprotection. TRPC channels are Ca(2+)-permeable, nonselective cation channels with a variety of physiological functions. Blocking TRPC channels with either a blocker or short-interfering RNAs (specific for TRPC 5 and 6) in primary neurons resulted in suppression of both PDGF-mediated neuroprotection as well as elevations in intracellular Ca(2+). PDGF-mediated neuroprotection involved parallel but distinct ERK/CREB and PI3K/Akt pathways. TRPC channel blocking also resulted in suppression of PDGF-induced Pyk2/ERK/CREB activation, but not Akt activation. Relevance of these findings in vivo was further corroborated by intrastriatal injections of PDGF and HIV-1 Tat in mice. Administration of PDGF was able to rescue the dopaminergic neurons in the substantia nigra from Tat-induced neurotoxicity. This effect was attenuated by pre-treatment of mice with the TRP blocker, thus underscoring the novel role of TRPC channels in the neuroprotection mediated by PDGF.
Figures









References
-
- Nath A, Anderson C, Jones M, Maragos W, Booze R, Mactutus C, et al. Neurotoxicity and dysfunction of dopaminergic systems associated with AIDS dementia. J Psychopharmacol. 2000;14(3):222–227. - PubMed
-
- Mocchetti I, Nosheny RL, Tanda G, Ren K, Meyer EM. Brain-derived neurotrophic factor prevents human immunodeficiency virus type 1 protein gp120 neurotoxicity in the rat nigrostriatal system. Ann N Y Acad Sci. 2007 Dec;1122:144–154. - PubMed
-
- Zauli G, Secchiero P, Rodella L, Gibellini D, Mirandola P, Mazzoni M, et al. HIV-1 Tat-mediated inhibition of the tyrosine hydroxylase gene expression in dopaminergic neuronal cells. J Biol Chem. 2000 Feb 11;275(6):4159–4165. - PubMed
-
- Eugenin EA, D'Aversa TG, Lopez L, Calderon TM, Berman JW. MCP-1 (CCL2) protects human neurons and astrocytes from NMDA or HIV-tat-induced apoptosis. J Neurochem. 2003 Jun;85(5):1299–1311. - PubMed
-
- Liu Y, Jones M, Hingtgen CM, Bu G, Laribee N, Tanzi RE, et al. Uptake of HIV-1 tat protein mediated by low-density lipoprotein receptor-related protein disrupts the neuronal metabolic balance of the receptor ligands. Nat Med. 2000 Dec;6(12):1380–1387. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
