

Mycobacterium tuberculosis WhiB3 maintains redox homeostasis by regulating virulence lipid anabolism to modulate macrophage response
- PMID: 19680450
- PMCID: PMC2718811
- DOI: 10.1371/journal.ppat.1000545
Mycobacterium tuberculosis WhiB3 maintains redox homeostasis by regulating virulence lipid anabolism to modulate macrophage response
Abstract
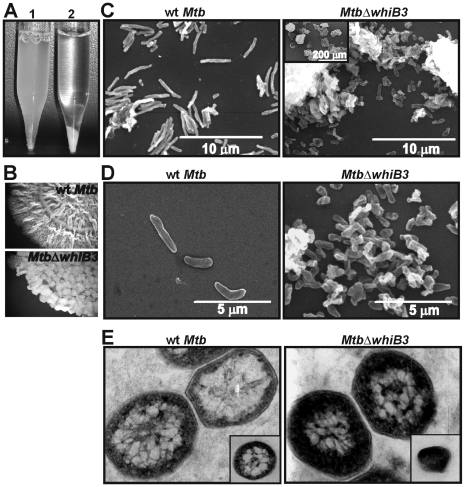
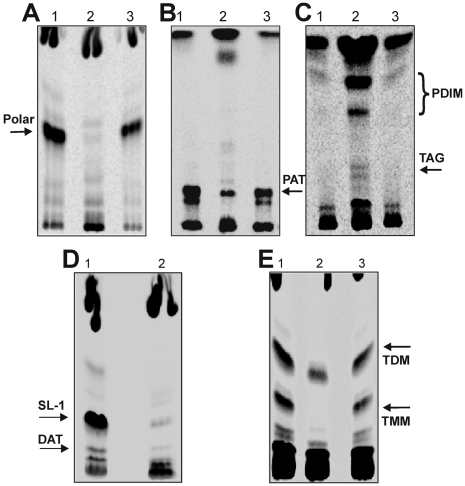
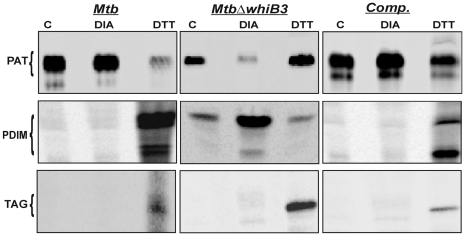
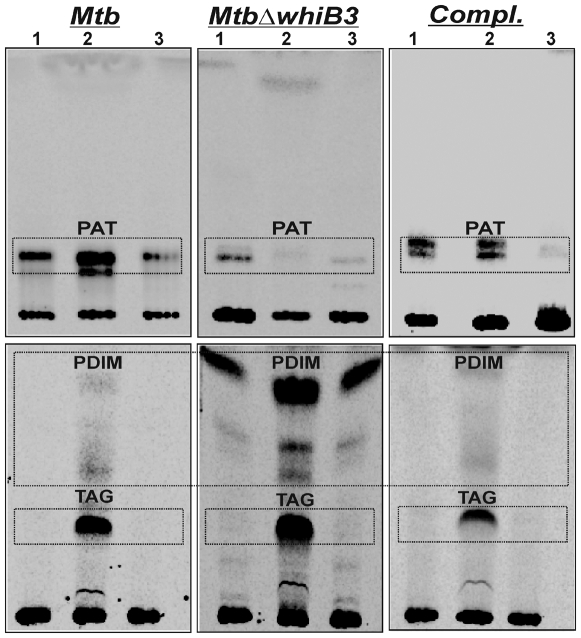
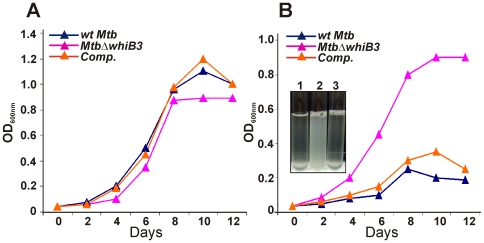
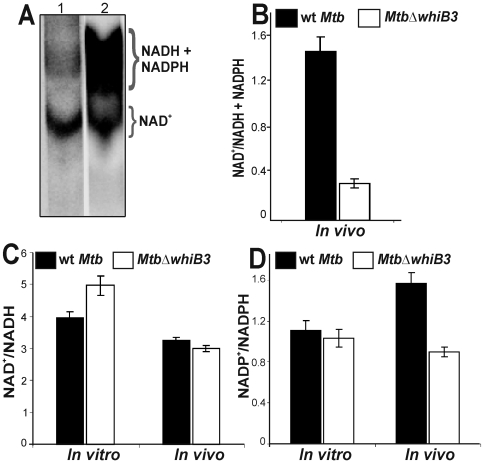
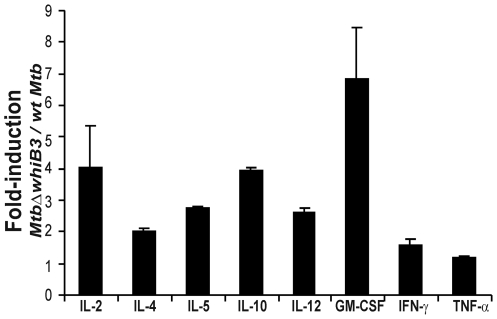
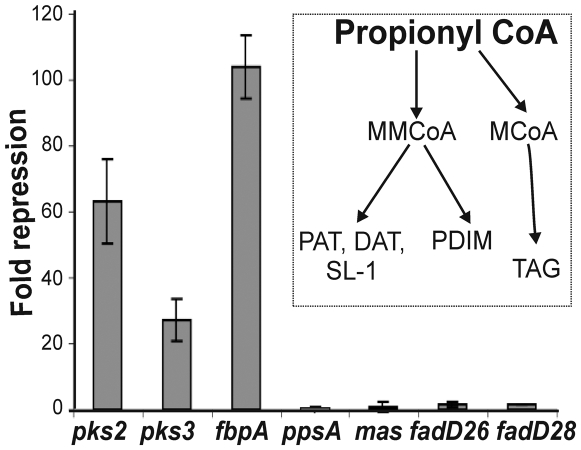
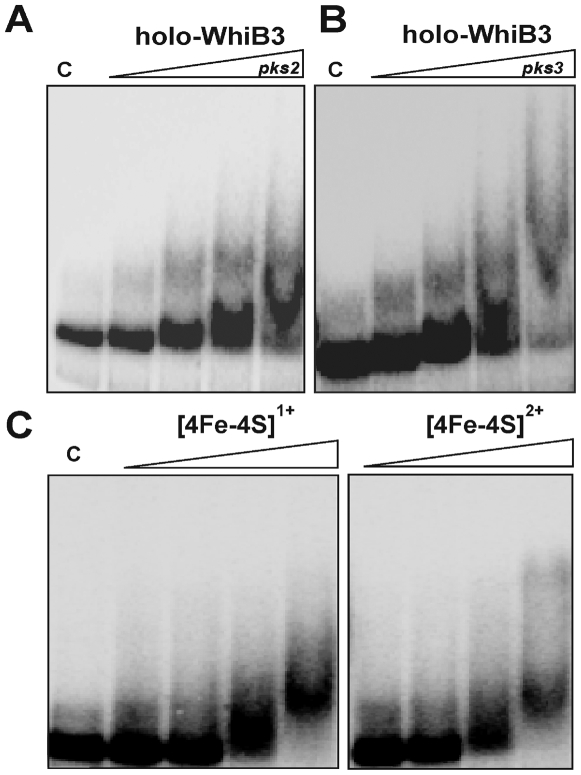
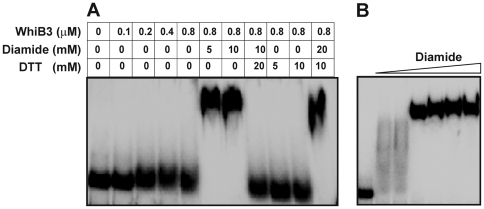
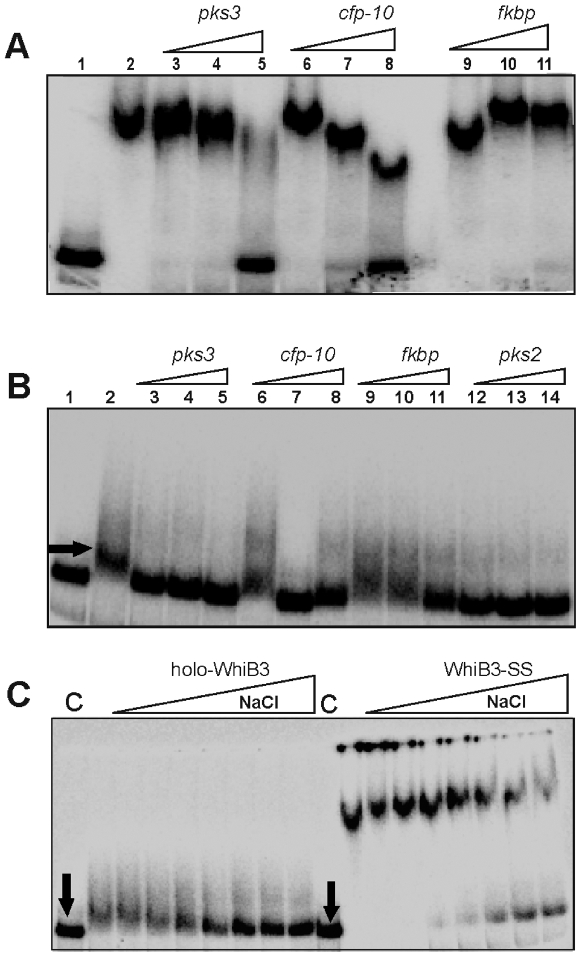
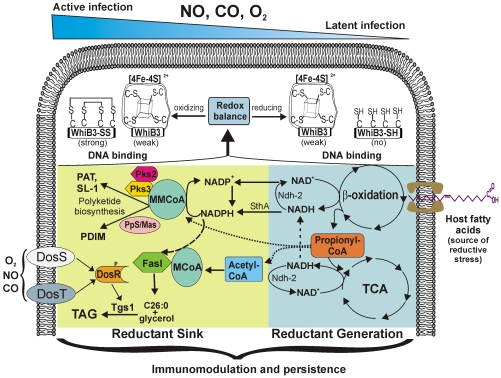
The metabolic events associated with maintaining redox homeostasis in Mycobacterium tuberculosis (Mtb) during infection are poorly understood. Here, we discovered a novel redox switching mechanism by which Mtb WhiB3 under defined oxidizing and reducing conditions differentially modulates the assimilation of propionate into the complex virulence polyketides polyacyltrehaloses (PAT), sulfolipids (SL-1), phthiocerol dimycocerosates (PDIM), and the storage lipid triacylglycerol (TAG) that is under control of the DosR/S/T dormancy system. We developed an in vivo radio-labeling technique and demonstrated for the first time the lipid profile changes of Mtb residing in macrophages, and identified WhiB3 as a physiological regulator of virulence lipid anabolism. Importantly, MtbDeltawhiB3 shows enhanced growth on medium containing toxic levels of propionate, thereby implicating WhiB3 in detoxifying excess propionate. Strikingly, the accumulation of reducing equivalents in MtbDeltawhiB3 isolated from macrophages suggests that WhiB3 maintains intracellular redox homeostasis upon infection, and that intrabacterial lipid anabolism functions as a reductant sink. MtbDeltawhiB3 infected macrophages produce higher levels of pro- and anti-inflammatory cytokines, indicating that WhiB3-mediated regulation of lipids is required for controlling the innate immune response. Lastly, WhiB3 binds to pks2 and pks3 promoter DNA independent of the presence or redox state of its [4Fe-4S] cluster. Interestingly, reduction of the apo-WhiB3 Cys thiols abolished DNA binding, whereas oxidation stimulated DNA binding. These results confirmed that WhiB3 DNA binding is reversibly regulated by a thiol-disulfide redox switch. These results introduce a new paradigmatic mechanism that describes how WhiB3 facilitates metabolic switching to fatty acids by regulating Mtb lipid anabolism in response to oxido-reductive stress associated with infection, for maintaining redox balance. The link between the WhiB3 virulence pathway and DosR/S/T signaling pathway conceptually advances our understanding of the metabolic adaptation and redox-based signaling events exploited by Mtb to maintain long-term persistence.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Dye C, Scheele S, Dolin P, Pathania V, Raviglione MC, et al. Global Burden of Tuberculosis: Estimated Incidence, Prevalence, and Mortality by Country. JAMA. 1999;282:677–686. - PubMed
-
- Reed MB, Domenech P, Manca C, Su H, Barczak AK, et al. A glycolipid of hypervirulent tuberculosis strains that inhibits the innate immune response. Nature. 2004;431:84–87. - PubMed
-
- Camacho LR, Constant P, Raynaud C, Laneelle MA, Triccas JA, et al. Analysis of the phthiocerol dimycocerosate locus of Mycobacterium tuberculosis. Evidence that this lipid is involved in the cell wall permeability barrier. J Biol Chem. 2001;276:19845–19854. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
