

Review
doi: 10.1016/j.mrfmmm.2009.08.002.
Epub 2009 Aug 12.
DNA polymerases at the eukaryotic fork-20 years later
Affiliations
- PMID: 19682465
- PMCID: PMC2822129
- DOI: 10.1016/j.mrfmmm.2009.08.002
Item in Clipboard
Review
DNA polymerases at the eukaryotic fork-20 years later
Mutat Res.
.
Abstract
Function of the eukaryotic genome depends on efficient and accurate replication of anti-parallel DNA strands. Eukaryotic DNA polymerases have different properties adapted to perform a wide spectrum of DNA transactions. Here we focus on major players in the bulk replication, DNA polymerases of the B-family. We review the organization of the replication fork in eukaryotes in a historical perspective, analyze contemporary models and propose a new integrative model of the fork.
Copyright (c) 2009 Elsevier B.V. All rights reserved.
Figures
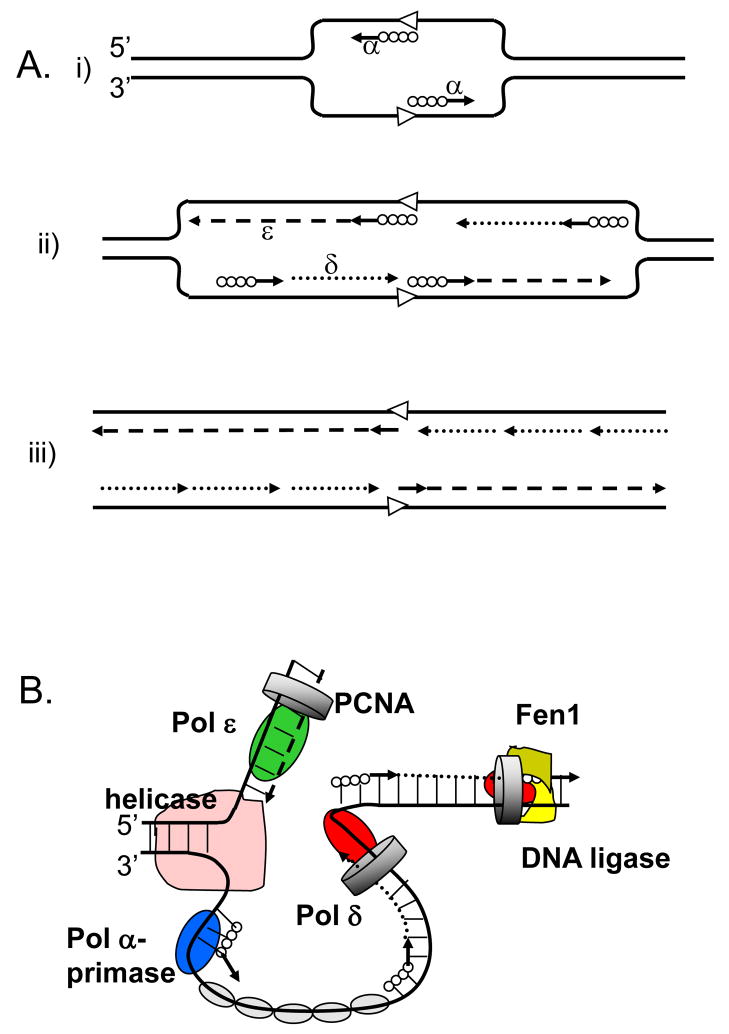
A. The model proposed in 1990 by Sugino group (adapted from [2]. (i) Pol α synthesizes short DNA segments (straight lines) primed by RNA (solid circles) at the replication origin (open arrowheads). (ii) Pol ε synthesizes the leading strand (dashed line); Pol α synthesizes short RNA-DNA stretches on the lagging strand, which are subsequently extended by Pol δ. (iii) After removal of RNA primers, Pol δ completes the lagging strand synthesis (dotted lines). B. The currently accepted model (adapted from [85]). The model illustrates primary roles for Pol ε (green oval) in leading DNA strand replication (dashed line) and Pol δ (red oval) in lagging strand replication (dotted line). Other proteins shown include the Pol α-primase (blue oval) synthesizing RNA-DNA hybrids (solid circles and straight lines), the MCM helicase (pink), the eukaryotic single-stranded-DNA-binding protein, replication protein A (RPA; gray ovals), the sliding clamp proliferating cell nuclear antigen (PCNA; gray ring) and the Fen1–DNA ligase complex (khaki-yellow).
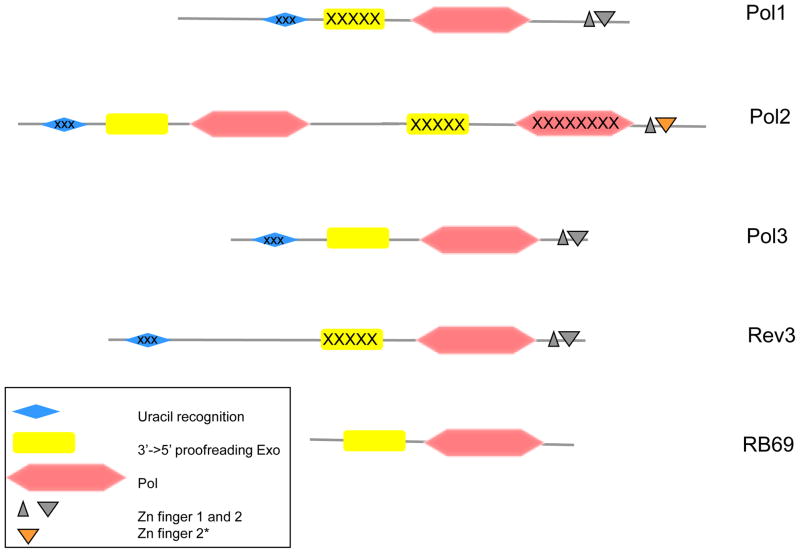
The drawing is based on the amino acid sequences of the catalytic subunits of yeast Pols and RB69 Pol and is roughly to scale. The inactivated C-terminal domains of Pol2, uracil-recognizing domains and exonuclease domains of Pol1 and Rev3 are marked by “X” symbols. Zn-finger 2* denotes the distinct version of this module in Pol2 that is highly similar to the Zn-finger of archaeal PolD [7].
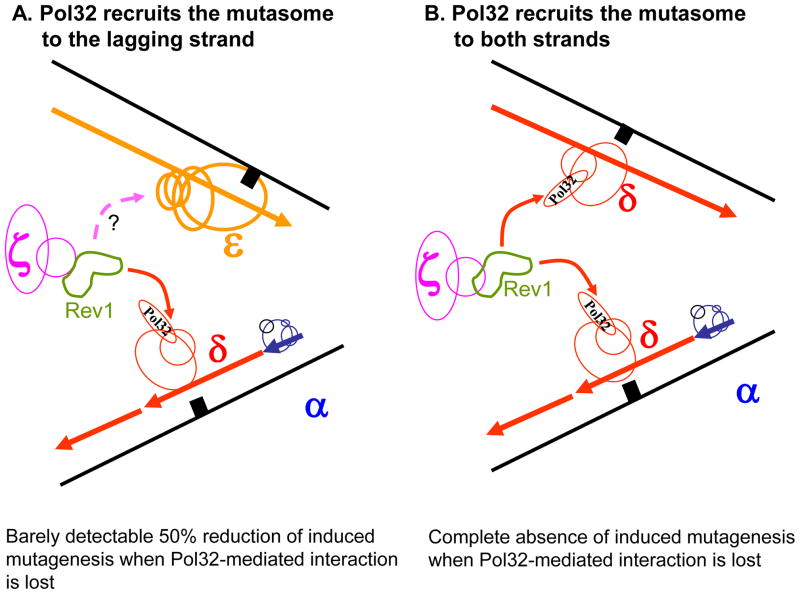
Pol32 mediates the recruitment of the mutasome including Rev1 and Pol ζ to the lagging strand (A) or both strands (B). Pol holoenzymes are drawn in blue (Pol α), red (Pol δ), yellow (Pol ε), purple (Pol ζ) and green (Rev1). A solid arrow acknowledges the proven interaction of Rev1 with Pol32. A broken arrow with a question mark indicates the hypothetical interaction of mutasome with Pol ε. A black square represents damaged DNA.
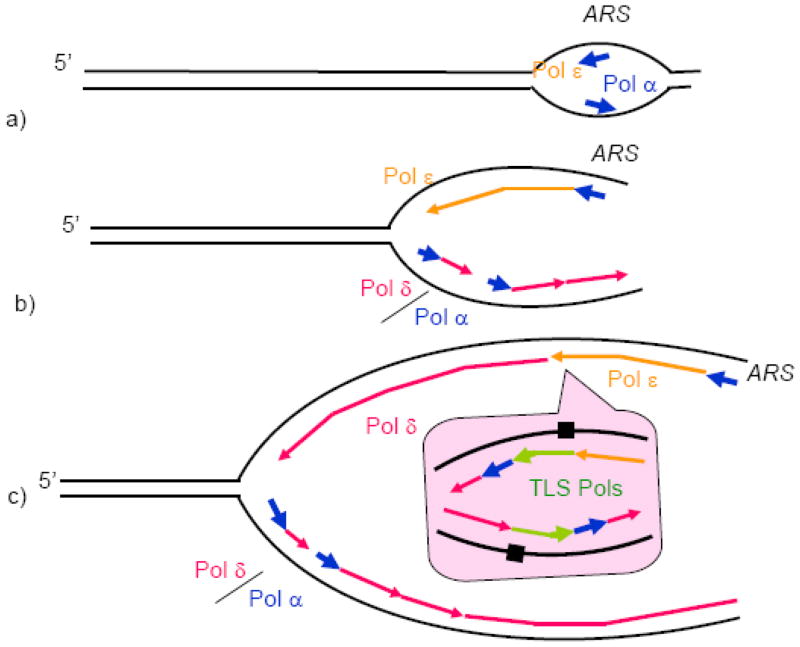
a) Pol ε and Pol α are recruited to the origins (ARS, standing for Autonomous Replication Sequence according to the yeast nomenclature) by the replication initiation machinery and start leading DNA strand synthesis. b) Pol δ is recruited to the lagging DNA strand, and replication of both strands proceeds further (only one fork moving to the left is shown for simplicity). c) At a random site away from the origin, Pol ε encounters an obstacle and dissociates. Its recruitment back is not possible due to the absence of origin-specific factors. Pol δ rapidly takes over the leading DNA strand synthesis. c, inset) Obstacles that can be overcome neither by Pol ε, nor by Pol δ, result in the recruitment of TLS polymerases. In the example shown, the replication block results in a replication restart downstream of the block, which requires Pol α and Pol δ. The remaining gaps are filled postreplicatively, as postulated by one of the current TLS models [98]. Replication block is shown as damaged DNA for simplicity. In reality, an elevated level of dissociation of the main replicative DNA polymerases due to genetic or physiological perturbations can results in the recruitment of TLS Pols to the fork.
References
-
- Morrison A, Araki H, Clark AB, Hamatake RK, Sugino A. A third essential DNA polymerase in S. cerevisiae. Cell. 1990;62:1143–1151. - PubMed
-
- Chilkova O, Stenlund P, Isoz I, Stith CM, Grabowski P, Lundstrom EB, Burgers PM, Johansson E. The eukaryotic leading and lagging strand DNA polymerases are loaded onto primer-ends via separate mechanisms but have comparable processivity in the presence of PCNANucleic. Acids Res. 2007;35:6588–6597. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
